| Issue |
Parasite
Volume 33, 2026
|
|
|---|---|---|
| Article Number | 28 | |
| Number of page(s) | 11 | |
| DOI | https://doi.org/10.1051/parasite/2026028 | |
| Published online | 19 May 2026 | |
Research Article
Phenology and parasitism rates of Xenos oxyodontes and Xenos moutoni (Strepsiptera: Xenidae) in Vespa (Hymenoptera: Vespidae) in South Korea
Phénologie et taux de parasitisme de Xenos oxyodontes et Xenos moutoni (Strepsiptera : Xenidae) chez Vespa (Hymenoptera : Vespidae) en Corée du Sud
1
Department of Applied Biology, Kyungpook National University, Daegu, Republic of Korea
2
Division of Forest Biodiversity, Korea National Arboretum, Pocheon, Republic of Korea
3
Institute of Agricultural Science and Technology, Kyungpook National University, Daegu, Republic of Korea
4
Department of Plant Medicine, College of Agriculture and Life Sciences, Kyungpook National University, Daegu, Republic of Korea
* Corresponding authors: This email address is being protected from spambots. You need JavaScript enabled to view it.
(Moon Bo Choi); This email address is being protected from spambots. You need JavaScript enabled to view it.
(Ohseok Kwon)
Received:
9
March
2026
Accepted:
24
April
2026
Abstract
The long-term seasonal parasitism patterns, hereafter referred to as stylopization, and within-nest developmental stage distributions of Vespa-associated Xenidae (Strepsiptera) remain poorly characterized in South Korea. We compiled 15 years of data (2008–2023) based on trapped Vespa individuals from multiple regions and quantified stylopization rates for two Xenidae species, Xenos oxyodontes and Xenos moutoni, based on diagnostic characters visible on stylopized hosts. Seasonal occurrence was summarized primarily from female Xenos specimens, which can be identified to species, and host-association patterns were assessed where possible. To complement the trap-based records, we dissected 22 Vespa analis colonies collected in September 2020 and documented strepsipteran developmental stages within the nests. Across 39,610 examined host individuals, stylopization rates and seasonal occurrence differed between the two Xenos species and among host taxa, indicating distinct host-use and temporal activity patterns. Importantly, the nest dissections provided direct observations of late-season developmental stage compositions, enabling us to compare within-nest reproductive/developmental stages to those inferred from trap-detected stylopized hosts. This comparison revealed that key reproductive-stage information may not be captured by trap records alone. Thus, integrating long-term trap data with nest-level observations can clarify species-specific phenology and developmental/reproductive stage distributions in Vespa-associated Xenidae, providing a better empirical context for interpreting stylopization intensity and developmental timing in field populations.
Résumé
Les schémas de parasitisme saisonnier à long terme, désignés ici par le terme de stylopisation, et la distribution des stades de développement au sein du nid des Xenidae (Strepsiptera) associés à Vespa restent mal connus en Corée du Sud. Nous avons compilé des données sur 15 ans (2008–2023) à partir d’individus de Vespa piégés dans plusieurs régions et quantifié les taux de stylopisation pour deux espèces de Xenidae, Xenos oxyodontes et Xenos moutoni, en nous basant sur les caractères diagnostiques visibles chez les hôtes stylopisés. L’occurrence saisonnière a été résumée principalement à partir de spécimens femelles de Xenos, qui peuvent être identifiés jusqu’à l’espèce, et les schémas d’association hôte-parasite ont été évalués lorsque cela était possible. Pour compléter les données de piégeage, nous avons disséqué 22 colonies de Vespa analis collectées en septembre 2020 et documenté les stades de développement des Strepsiptères au sein des nids. Sur 39 610 individus hôtes examinés, les taux de stylopisation et la fréquence saisonnière différaient entre les deux espèces de Xenidae et entre les taxons hôtes, indiquant des profils d’utilisation de l’hôte et d’activité temporelle distincts. Surtout, la dissection des nids a fourni des observations directes de la composition des stades de développement en fin de saison, nous permettant de comparer les stades de reproduction et de développement au sein des nids à ceux déduits des hôtes stylopisés détectés par piégeage. Cette comparaison a révélé que des informations clés sur les stades de reproduction ne sont pas nécessairement accessibles par les seuls relevés de piégeage. Ainsi, l’intégration de données de piégeage à long terme avec des observations au niveau des nids peut clarifier la phénologie spécifique à l’espèce et la distribution des stades de développement et de reproduction chez les Xenidae associés à Vespa, fournissant un meilleur contexte empirique pour interpréter l’intensité de la stylopisation et le calendrier de développement dans les populations naturelles.
Key words: Host association / Voltinism / Developmental stage / Parasitism
Edited by Jean-Lou Justine
© J. Kim et al., published by EDP Sciences, 2026
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
Species of the order Strepsiptera are obligate endoparasites of other insects, and they are well known for their extreme sexual dimorphism and highly specialized reproductive biology [12, 13, 17, 31]. In all but the primitive members of Mengenillidae, females exhibit neoteny and remain larviform within the host, where they act as parasites and reproduce, whereas males emerge, becoming free-living adults, and are typically short-lived. After reaching maturity inside the host, the female exposes only the cephalothorax through the host’s abdominal integument, and mating occurs at this exposed region. The female then larviposits triungulins. These triungulins represent the dispersal stage; they leave the original host, move to a new host, and enter its body, where development continues [2, 8, 14, 16, 28, 31]. Given this life history, strepsipteran parasitism is best viewed as a long-term interaction that is tightly coupled to the host’s survival and seasonal activity.
Specialized strepsipteran parasitism is commonly referred to as stylopization, and a wide range of physiological and behavioral alterations have been documented in stylopized hosts [1, 2]. Such effects have been discussed with particular frequency for species in the strepsipteran family Xenidae, which parasitize eusocial vespid wasps. Among vespids, the Polistes–Xenos association is the most well-studied, with reported effects including host sterilization, the suppression of ovarian development, the cessation of worker tasks, nest abandonment, and extranidal aggregation [9, 10, 18, 27]. Transcriptomic evidence further suggests that in Polistes, stylopization can be accompanied by changes in gene-expression patterns in the brain [2, 6], raising the possibility that transcriptomic profiles associated with the worker caste are attenuated and remodeled toward a more gyne-like state. Such shifts also appear to be associated with extended worker longevity. Mature strepsipteran females can induce diapause in workers after mating [4, 34], and larviposition has been reported to occur the following spring after the host terminates diapause [23, 25, 30, 34].
Outside of the Polistes–Xenos system, basic ecological information on many Vespa–Xenos associations, including host specificity and seasonal developmental patterns, remains relatively limited, despite the existence of observational records. Among Vespa-associated Xenos species, Xenos oxyodontes Nakase & Kato, 2013 and Xenos moutoni Buysson, 1903 are the most commonly referenced. They were previously treated as the same species, X. moutoni [22, 24], but were recently redescribed as distinct cryptic species [29]. The two species are generally considered to differ in host specificity. Stylopization by X. oxyodontes has primarily been recorded in Vespa analis Fabricius, 1775, with additional cases reported from Vespa simillima Smith, 1868 and Vespa velutina de Lepeletier, 1836, whereas stylopization by X. moutoni has been observed in multiple Vespa species, but not V. analis [11, 15, 29]. This distinction suggests that species-level resolution within Xenos is necessary when interpreting phenology and parasitism ecology. In Japan, studies have quantified seasonal occurrence patterns and parasitism levels for X. moutoni and X. oxyodontes, providing an important baseline for interpreting field observations of Vespa-associated Xenos species [21, 23, 25, 30]. However, while trap-based data are useful for describing seasonal occurrence, they cannot directly show where individuals transition into the developmental and reproductive stages. Thus, empirical data regarding whether Vespa-associated Xenos mate within or outside of the host’s nest are scarce. To acquire such data, nest dissection can be used to characterize patterns of mature parasite stages within the nest and to formulate hypotheses on the spatial context of reproductive events [23].
Our study aimed to provide a baseline synthesis of the phenology and stylopization patterns of two Xenidae species, X. moutoni and X. oxyodontes, in Vespa using material collected in South Korea. First, using multi-year, multi-region trap data, we examined seasonal occurrence patterns and geographic distributions based primarily on female specimens, which can readily be identified to species, and we also assessed host-association patterns where possible. In addition, to evaluate the distribution of developmental and reproductive stages of Xenos, information not readily inferable from trap data alone, we used stylopized hosts collected from V. analis nests to characterize the reproductive-stage distributions of female and male X. oxyodontes individuals within hornet colonies. By integrating the broad-scale temporal patterns provided by trap-based records with the stage-specific information obtained from colony samples, we refined the baseline knowledge on when, where, and during which stages stylopization occurs in hornets, and, within the limits of our data, we make conservative inferences regarding expected local stylopization intensities and voltinism.
Materials and methods
Specimen collection
Nest collection
On September 2, 2020, we collected 22 active V. analis colonies from Muui Island (Jung-gu, Incheon Metropolitan City, South Korea). For each colony, recovered individuals were preserved in 99% ethanol, and we recorded the number of individuals by caste (queen, worker, and male).
Trapping
Trap surveys were conducted from 2008 to 2023. In Incheon and Gyeonggi-do, traps were fixed at the same sites for multiple years, with additional traps installed in other regions during specific years. A total of 677 traps comprising a mixture of hornet attractant traps and Malaise traps were deployed (Fig. 1, Supplementary file). Traps were installed each year in May, and insects were retrieved at approximately one-month intervals until trap removal in November, and a single “visit,” as used in the statistical analyses, was defined as one trap-check and collection event. From the collected samples, Vespa individuals were sorted and preserved in 99% ethanol.
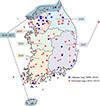 |
Figure 1 Spatial distribution of 677 trapping sites in South Korea (2008–2023). Blue squares represent Malaise trap sites, sampled from 2008 to 2018, and red circles indicate sugar-attractant trap sites, sampled from 2019 to 2023. Shaded areas approximate the year-specific sugar attractant trap survey coverage over the 2019–2023 period, with the years specified on the map. |
Specimen examination and classification
All specimens were examined under a stereomicroscope, and individuals with a strepsipteran cephalothorax protruding between the abdominal tergites were confirmed as stylopized hosts (Fig. 2). Stylopized hornets were stored individually, and we recorded Vespa species and caste and Xenos species and sex (based on the female cephalothorax or traces of male puparium/adult emergence). Female caste was distinguished based on relative body size [27]. Xenos were identified following Nakase and Kato (2013) [29].
 |
Figure 2 A female Xenos moutoni stylopizing a host hornet, Vespa crabro, protruding from the host abdomen. |
Xenos individuals were assigned to stages based on sex-specific reproductive characteristics: females were classified as pre-vitellogenic, vitellogenic, gravid, larvipositing, or post-larviposition, and males were classified as pupa, transitional, not emerged, or emerged (Table 1). For trap samples, since installation and collection dates differed among traps, we used the midpoint date between trap installation and collection as the capture timing for each specimen to minimize error.
Diagnostic criteria for reproductive and developmental stage assignment in Xenos spp.
Statistical analysis
All statistical analyses and visualizations were conducted in R (4.4.2).
Phenology analysis
To evaluate differences in female Xenos phenology among reproductive stages, we compared the distributions of capture times (midpoint date) across stages. Because the data did not meet the assumption of normality and sample sizes among stages were unbalanced, we used a Kruskal–Wallis test. When a significant difference was detected, we conducted post hoc pairwise comparisons using Dunn’s test with the Holm correction for multiple comparisons. We calculated the effect size (ε2) as an index of the extent to which stage membership explains variation in midpoint capture time:
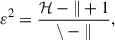
H: test statistic
k: number of groups
To maintain consistency in sample sizes among stages, we pooled pre-vitellogenic and vitellogenic females into a single group and pooled gravid and larvipositing females into a single group, yielding three reproductive-stage groups for day of year (DOY) distribution comparisons.
Distributional analysis
Trap-based surveys are subject to spatial capture bias, such that in some areas, species may be present but, nevertheless, go undetected [32]. For eusocial wasps, in particular, attraction to sugar-based baits varies with bait type and resource responsiveness, and trap efficiency can therefore differ markedly across species and environmental contexts [20, 26]. In our analysis, the response variable was defined as the number of stylopized hosts per trap visit for each trap (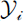 ). To estimate the trap-level intensity of Xenos stylopization, we fitted a negative binomial generalized linear model to
). To estimate the trap-level intensity of Xenos stylopization, we fitted a negative binomial generalized linear model to  using a log link, including the total number of host individuals captured during the same trap visit (Hᵢ) as an offset. This analysis follows Heusler et al. (2025) [7] and is not intended to infer the causal effects of environmental covariates, but to summarize where and to what extent host-capture–adjusted expected stylopized-host events are spatially concentrated within a consistent spatial sampling framework.
using a log link, including the total number of host individuals captured during the same trap visit (Hᵢ) as an offset. This analysis follows Heusler et al. (2025) [7] and is not intended to infer the causal effects of environmental covariates, but to summarize where and to what extent host-capture–adjusted expected stylopized-host events are spatially concentrated within a consistent spatial sampling framework.
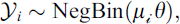
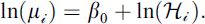
Here, 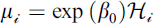 represents the expected number of stylopization events, with
represents the expected number of stylopization events, with 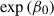 corresponding to the mean stylopization event rate per host individual; θ is the overdispersion parameter of the negative binomial distribution; and Hi denotes the number of potential host individuals captured during the same trap visit, with in
corresponding to the mean stylopization event rate per host individual; θ is the overdispersion parameter of the negative binomial distribution; and Hi denotes the number of potential host individuals captured during the same trap visit, with in 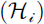 included as an offset to adjust for differences in host captures among trap visits. We used the estimated μi value as the expected stylopized host count for each trap.
included as an offset to adjust for differences in host captures among trap visits. We used the estimated μi value as the expected stylopized host count for each trap.
For the spatial analysis of X. moutoni, we included all traps in which hornets were captured as analysis units, reflecting the strepsipteran species’ relatively broad host spectrum among Vespa species. In contrast, X. oxyodontes is more host-specific, and traps without captures of its primary hosts, V. analis and V. simillima simillima, provide no opportunity for stylopization estimation. Accordingly, for the spatial analysis of X. oxyodontes, we restricted the analysis to traps that captured at least one V. analis or mainland V. simillima individual, excluding structural zeros arising from host absence. Therefore, the results should be interpreted as variation in stylopization intensity conditional on host detection. Although X. oxyodontes has occasionally been reported from V. velutina, no stylopized individuals were detected in this host in our trap samples. Therefore, traps capturing only V. velutina were not included as potential host records in the distribution analysis.
To visualize spatial patterns, we applied hexagonal binning based on trap coordinates. Within each hexagon, we calculated the mean of 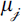 across all visits to traps contained within that cell (
across all visits to traps contained within that cell (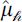 ) to obtain a spatially aggregated expected stylopized-host count. This aggregation was used to smooth local variability at the individual-trap level and to summarize broad-scale spatial patterns. To compare the spatial concentrations of
) to obtain a spatially aggregated expected stylopized-host count. This aggregation was used to smooth local variability at the individual-trap level and to summarize broad-scale spatial patterns. To compare the spatial concentrations of 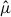 values between X. moutoni and X. oxyodontes, we ranked hexagons in descending order of
values between X. moutoni and X. oxyodontes, we ranked hexagons in descending order of 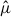 and then calculated the cumulative proportion of
and then calculated the cumulative proportion of 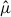 represented by each species as a function of the cumulative proportion of hexagons, constructing rank–cumulative curves. Concentration was quantified in two ways, as the proportion of the total cumulative
represented by each species as a function of the cumulative proportion of hexagons, constructing rank–cumulative curves. Concentration was quantified in two ways, as the proportion of the total cumulative 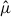 values accounted for by the top 10% of the hexagons, and by the area under the rank–cumulative curve (AUC). We defined AUC as a concentration index that quantifies deviation of the rank–cumulative curve from the uniform reference line (the diagonal) and interpreted larger AUC values as indicating a stronger concentration of
values accounted for by the top 10% of the hexagons, and by the area under the rank–cumulative curve (AUC). We defined AUC as a concentration index that quantifies deviation of the rank–cumulative curve from the uniform reference line (the diagonal) and interpreted larger AUC values as indicating a stronger concentration of 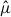 values in a smaller number of hexagons.
values in a smaller number of hexagons.
To assess whether the observed concentration metrics deviated from a random distribution, we conducted a permutation test. For each species, we randomly reassigned the 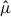 values across hexagon IDs and recalculated the concentration metrics, repeating this procedure 10,000 times to generate a null distribution. The p-value was then computed as the proportion of permutation runs in which the concentration metric was equal to or more extreme than the observed value.
values across hexagon IDs and recalculated the concentration metrics, repeating this procedure 10,000 times to generate a null distribution. The p-value was then computed as the proportion of permutation runs in which the concentration metric was equal to or more extreme than the observed value.
Because our model included only an intercept and host availability, the spatial pattern of 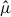 primarily reflects how host captures are distributed within the sampling frame rather than a spatial effect of the parasite itself. Accordingly, we interpreted the results of the analysis described in this section as the spatial concentration of expected stylopization events under a consistent survey design, rather than as spatial heterogeneity in parasitism rate. Since no stylopization was observed in Vespa individuals captured in the two major island regions, Jeju Island and Ulleung Island (Table 2), these records were excluded from the distribution analysis.
primarily reflects how host captures are distributed within the sampling frame rather than a spatial effect of the parasite itself. Accordingly, we interpreted the results of the analysis described in this section as the spatial concentration of expected stylopization events under a consistent survey design, rather than as spatial heterogeneity in parasitism rate. Since no stylopization was observed in Vespa individuals captured in the two major island regions, Jeju Island and Ulleung Island (Table 2), these records were excluded from the distribution analysis.
Number and stylopization status of Vespa individuals collected in traps across South Korea over fifteen years, organized by Vespa and Xenos species.
Results
A total of 39,610 individuals representing eight host species were collected, of which 181 were confirmed as stylopized hosts (Table 2). Among host species, Vespa crabro was the most frequently collected (9,748 individuals), whereas Vespa dybowskii was the least collected (1,726 individuals), and no stylopized individuals were detected for this species (0%). Additionally, no stylopized V. velutina individuals were detected, despite 4,302 individuals being collected (0%). The largest number of stylopized hosts was recorded for V. analis (58 individuals; 3.320%), followed by V. crabro (50 individuals; 0.513%), and Vespa mandarinia (36 individuals; 1.846%). The lowest rate was observed for V. simillima (Mainland), in which X. oxyodontes was detected at 0.022%. Notably, females in the larviposition stage were recorded in trap captures during July–September and also in October, and many of these records were from worker hosts. No stylopization was observed in Vespa captured on Jeju Island and Ulleung Island, the two major island regions.
Phenology
In X. moutoni, no significant difference among stages was detected (H = 0.405, df = 2, p = 0.817) (Fig. 3A). The pre-vitellogenic/vitellogenic stage had a median DOY of 230 (IQR of 220–253), the gravid/larvipositing stage had a median DOY of 224 (IQR of 217–244), and the post-larviposition stage had a median DOY of 224 (IQR of 217–240). Since stage differences were not significant in the Kruskal–Wallis test, we did not further interpret stage-specific differences for X. moutoni.
 |
Figure 3 Monthly life-stage compositions of female (A) Xenos moutoni and (B) Xenos oxyodontes individuals captured by traps in South Korea. The lines (top) indicate the monthly number of captured females (n), and the stacked bars (bottom) show the within-month proportional compositions of female reproductive stages. |
In contrast, significant differences among stages were detected in X. oxyodontes (H = 12.6, df = 2, p = 0.00182) (Fig. 3B). Dunn’s post hoc test with the Holm adjustment indicated that the pre-vitellogenic/vitellogenic and gravid/larvipositing stages were significantly different (p_adj = 0.00162), whereas other pairwise comparisons were not. The effect size for X. oxyodontes was ε2 = 0.342, indicating stage-associated differences in capture timing. The pre-vitellogenic/vitellogenic stage had a median DOY of 284 (IQR of 262–294), the gravid/larvipositing stage had a median DOY of 224 (IQR of 214–239), and the post-larviposition stage had a median DOY of 244 (IQR of 240–248). For X. oxyodontes, individuals in the gravid/larvipositing stage tended to be captured earlier in the year than those in the pre-vitellogenic/vitellogenic stage, and the overall effect of stage was primarily driven by the separation between these two stages.
Distribution
To analyze the distributions of both Xenos species, we characterized the spatial concentrations of host-capture–adjusted expected stylopized-host counts (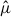 ) by applying a consistent sampling frame and fitting a negative binomial GLM with an offset for host captures to the trap data. High
) by applying a consistent sampling frame and fitting a negative binomial GLM with an offset for host captures to the trap data. High 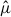 -value hexagons tended to be more locally concentrated within the sampling frame for X. oxyodontes than for X. moutoni (Fig. 4). The top 10% of hexagons accounted for 23.94% of the total expected stylopized-host counts in X. moutoni, whereas the top 10% of hexagons accounted for 58.08% in X. oxyodontes (Figs. 4A, 4B), and the rank-curve–based concentration metric (AUC) was higher in X. oxyodontes (AUC = 0.827 for X. oxyodontes vs AUC = 0.736 for X. moutoni), further supporting our finding that this species’ expected stylopization events are more locally concentrated. Permutation tests indicated that the observed concentration metrics deviated significantly from those of the random-reassignment distributions (Fig. 4C). Of note, because the spatial summary for X. oxyodontes is conditional on trap visits after excluding structural zeros (i.e., traps in which the primary hosts were not observed), this finding should be interpreted as the concentration of expected events within the sampling frame given host detection.
-value hexagons tended to be more locally concentrated within the sampling frame for X. oxyodontes than for X. moutoni (Fig. 4). The top 10% of hexagons accounted for 23.94% of the total expected stylopized-host counts in X. moutoni, whereas the top 10% of hexagons accounted for 58.08% in X. oxyodontes (Figs. 4A, 4B), and the rank-curve–based concentration metric (AUC) was higher in X. oxyodontes (AUC = 0.827 for X. oxyodontes vs AUC = 0.736 for X. moutoni), further supporting our finding that this species’ expected stylopization events are more locally concentrated. Permutation tests indicated that the observed concentration metrics deviated significantly from those of the random-reassignment distributions (Fig. 4C). Of note, because the spatial summary for X. oxyodontes is conditional on trap visits after excluding structural zeros (i.e., traps in which the primary hosts were not observed), this finding should be interpreted as the concentration of expected events within the sampling frame given host detection.
 |
Figure 4 Spatial distributions of the GLM-based, host-capture–adjusted expected stylopized-host counts ( |
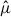
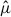
Nest-level patterns
Among 22 V. analis nests collected on Muui-do, Incheon (Table 3, Fig. 5), a total of 2,155 hornet individuals were recorded, of which 75 were parasitized. The mean number of hornet individuals per nest was approximately 98 (2,155/22), and the mean number of parasitized individuals per nest was approximately 3.41 (75/22). Nest-level stylopization rates showed substantial variation, ranging from 0% to 15.385%. The highest stylopization rate was observed in Nest 14 (15.385%), followed by Nest 22 (12.000%). In contrast, seven nests contained no stylopized individuals.
Colony composition and stylopization in 22 V. analis nests collected from Muui-do, Incheon, South Korea on September 2, 2020.
 |
Figure 5 (A) Number of V. analis individuals stylopized by X. oxyodontes in each of the 22 colonies examined. Dark grey bars represent females, and white bars represent males. (B) Colony-level stylopization rate of V. analis by X. oxyodontes, calculated as the proportion of stylopized individuals among all adults examined in each nest. Each dot represents one V. analis colony, and the raincloud plot shows the distribution of colony-level stylopization rates. |
Across the 22 nests, the occurrence of stylopized hornets showed substantial among-nest variation, with no stylopized hornet detected in 31.82% of the nests, whereas stylopized hornets were confirmed in many (Fig. 5). A total of 79 X. oxyodontes individuals were recorded, comprising 28 females and 51 males (Fig. 6). Regarding developmental stages, Xenos males were mostly in puparium-associated stages (pupa, transitional, or not emerged), and no evidence of puparium emergence was observed. Females were in either the pre-vitellogenic or vitellogenic stage, whereas individuals in the gravid, larviposition, or post-larviposition stages were not detected (Figs. 6 and 7).
 |
Figure 6 Sex-specific developmental stage composition of X. oxyodontes in stylopized V. analis individuals collected from 22 nests. |
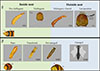 |
Figure 7 Representative developmental stages of Xenos oxyodontes recovered from inside and outside Vespa analis nests. Vitellogenic females and not-emerged males were detected in both inside- and outside-nest samples, suggesting that nest departure may occur at or around these stages. Abbreviations: WB, whole body (female); Co, cephalothorax (female); HA, host abdomen; EP, empty puparium (male); P, puparium (male); WBP, whole body in puparium (male). |
Discussion
This study combined 15 years of trap data with nest dissection data from South Korea. We found that the seasonal timing of occurrence differed among reproductive stages in female X. oxyodontes, whereas stage-specific differences were not evident in X. moutoni, and that high expected stylopized-host counts per sampling frame tended to be more locally concentrated for X. oxyodontes than for X. moutoni. In the V. analis nests, mature reproductive stages were not detected among Xenos, with only pre-reproductive stages observed.
Host range and detection probability
Our trap results showed that expected stylopization tended to be spatially non-uniform for both Xenos species, with stronger local concentrations for X. oxyodontes than for X. moutoni, suggesting that stylopization by either species may be greater in specific regions or amplified under certain conditions. However, such concentration could be exaggerated by differences in host availability, habitat- or host-use patterns, or trap-based detection probabilities, and should therefore be validated by pairing nest-based sampling with trap data within the same localities [11]. Although nest-based surveys have reported that V. velutina, like V. simillima, can be stylopized by both X. oxyodontes and X. moutoni [15], no stylopized V. velutina individuals were detected in our trap samples, suggesting that differences between nest-based and trap-based sampling frameworks strongly influence observed patterns [11].
Stage-specific phenology and extranidal emergence
The time of year in which different reproductive stages of female X. oxyodontes were encountered in trapped Vespa wasps differed. In contrast, X. moutoni showed no clear stage-specific seasonal differences, with stages occurring in a relatively uniform pattern. This may suggest that transitions among developmental stages in X. oxyodontes are concentrated within specific seasonal periods. Larvipositing-stage female X. oxyodontes individuals found in the host during late autumn would be expected to have invaded host eggs produced within the same year, suggesting these individuals may represent a second or later generation, arising from parasitism that occurred during the midseason. Interestingly, a phenology study on this species in Japan proposed that individuals detected in nests during the late season represent the second generation of the year, and the authors argued for a bivoltine cycle in this species [23]. Similar logic has been considered for the European species Xenos vesparum Rossi, 1793, for which Beani et al. (2018) [3] reported a bivoltine cycle in Polistes wasps in Italy, with female parasites releasing larvae twice, once in spring and once in summer. Therefore, the seasonal distribution of X. oxyodontes in South Korea should also be interpreted with the possibility of a bivoltine or multivoltine life cycle in mind [19].
The absence of evidence for male emergence and the lack of reproductive stage females in our nest samples are consistent with the possibility that some key reproductive events occur outside the nest. Importantly, in V. analis wasps collected using traps, male puparia have been reported to be largely absent, raising the possibility that hosts leave the nest before male emergence, such that emergence occurs outside the nest. Similarly, Makino (2001) [21] hypothesized, based on trap-based observations, that mating may not be restricted to the interior of the host’s nest. Strepsipterans have a life history in which the female remains within the host’s body, and because dispersal to new hosts depends on triungulin transfer, host movements are repeatedly discussed as being linked to transmission success [12, 13]. In the Polistes–Xenos association system in particular, field observations and experiments have shown that stylopized hosts cease colony tasks, leave the nest, and aggregate externally, and it is believed that these aggregation sites may coincide with mating, larval release, and overwintering [2, 10]. As males are extremely short-lived, it is especially important that females attract them via pheromones during their brief lifespan, and reproductive maturity is, thus, expected to be synchronized between the sexes [5, 8, 12]. This context makes our findings even more intriguing, raising further questions about the timing of mating in X. oxyodontes. However, because our nest dissections were based on a single early-September time point, the present data alone cannot distinguish between whether mature individuals had already departed or simply whether pre-mature stages predominated at the time of sampling.
While late-season larviposition was documented in our trap records, no mature reproductive stages were detected in the early-September nest dissections, making it difficult to conclude that the life cycle is completed entirely within the host’s nest. Although triungulins are often treated as a key element in discussions of voltinism, it remains necessary to determine whether triungulins released in the September–October period can proceed through overwintering. Notably, prior empirical studies have repeatedly supported the idea that stylopization prolongs host survival, allowing stylopized hosts to survive into the following year, and the strepsipteran subsequently larviposits [4, 33]. Accordingly, our data are not simply a reiteration of an extranidal scenario; the central implication is that there is a need to determine which pathways, such as host overwintering and extended longevity, allow late-season larviposition or triungulin release to connect to the next season, i.e., what mechanisms link successive generations [21].
For X. moutoni, however, given that worker-caste hosts are unlikely to survive for extended periods in the way queens do, it is difficult to explain how a cohort produced in late-season (September–October) larviposition could overwinter successfully and resume activity the following spring. The observation of larvipositing-stage females in September–October, and the fact that these cases were primarily associated with worker hosts, suggests that – given the known constraints on the worker lifespan – larvae released within the same year may have the potential to carry over into the next generation. Thus, a multivoltine life history may be common in Korean populations, but because our study does not include direct individual tracking, we present bi- or multi-voltinism only conservatively here, at the level of plausibility. This line of inference could be strengthened in future work by combining concurrent sampling at extranidal aggregation and overwintering sites with season-long series of nest samples.
Conclusion
In this study, we combined 15 years of trap data from South Korea with dissections of V. analis nests to provide a baseline synthesis of the phenology and stylopization patterns of two xenids, X. oxyodontes and X. moutoni. In the trap-based analyses, X. oxyodontes showed a tendency for the occurrence of female reproductive stages to vary seasonally, whereas stage-specific differences were limited in X. moutoni, and the spatial distributions of the two species were not equivalent within the shared sampling frame. Specifically, the concentration of high host-capture–adjusted expected stylopized-host counts was more localized in X. oxyodontes than in X. moutoni. In contrast, no mature reproductive-stage Xenos individuals were collected from V. analis nests in an early September sampling, with only pre-mature stages found, raising the possibility that some key reproductive events occur outside the nest or that late-season larviposition detected in trapped hosts and within-nest stage compositions may be temporally and/or spatially decoupled. Accordingly, rather than asserting voltinism, our data sharpen the research agenda by highlighting the need for a better understanding of how late-season larviposition and triungulin release are linked to the subsequent season. Future work that couples trap and nest sampling within the same localities and incorporates season-long series of nest samples could elucidate this link.
Acknowledgments
This article is based on the Master’s thesis of Jaehee Kim, completed at the Department of Applied Biology, Kyungpook National University. We also thank the Korea National Arboretum for providing parasitic specimen samples.
Funding
This work was supported by grants from the National Research Foundation of Korea (NRF), funded by the Korean Government (MSIT) (No. RS-2023-00212908)
Conflicts of interest
The authors declare that they have no conflict of interest.
Supplementary material
Supplementary file: Geographic coordinates and annual deployment information for the 677 trap sites used in this study. Access Supplementary Material
References
- Baudoin M. 1975. Host castration as a parasitic strategy. Evolution, 29, 335–352. [Google Scholar]
- Beani L. 2006. Crazy wasps: when parasites manipulate the Polistes phenotype. Annales Zoologici Fennici, 43, 564–574. [Google Scholar]
- Beani L, Cappa F, Manfredini F, Zaccaroni M. 2018. Preference of Polistes dominula wasps for trumpet creepers when infected by Xenos vesparum: a novel example of co-evolved traits between host and parasite. PLoS One, 13, e0205201. [Google Scholar]
- Beani L, Dallai R, Cappa F, Manfredini F, Zaccaroni M, Lorenzi MC, Mercati D. 2021. A strepsipteran parasite extends the lifespan of workers in a social wasp. Scientific Reports, 11, 7235. [CrossRef] [PubMed] [Google Scholar]
- Cvačka J, Jiroš P, Kalinová B, Straka J, Černá K, Šebesta P, Tomčala A, Vašíčková S, Jahn U, Šobotník J. 2012. Stylopsal: the first identified female-produced sex pheromone of Strepsiptera. Journal of Chemical Ecology, 38, 1483–1491. [Google Scholar]
- Geffre AC, Liu R, Manfredini F, Beani L, Kathirithamby J, Grozinger CM, Toth AL. 2017. Transcriptomics of an extended phenotype: parasite manipulation of wasp social behaviour shifts expression of caste-related genes. Proceedings of the Royal Society B: Biological Sciences, 284, 20170029. [Google Scholar]
- Heusler J, Funk J, Wagner A. 2025. Spatial interpolation in applied insect ecology: a review, including guidelines and datasets for practical use. Journal of Applied Entomology, 149, 1319–1334. [Google Scholar]
- Hrabar M, Danci A, McCann S, Schaefer PW, Gries G. 2014. New findings on life history traits of Xenos peckii (Strepsiptera: Xenidae). Canadian Entomologist, 146, 514–527. [CrossRef] [Google Scholar]
- Hughes D. 2002. The value of a broad mind: some natural history meanderings of Bill Hamilton. Ethology Ecology & Evolution, 14, 83–89. [Google Scholar]
- Hughes DP, Kathirithamby J, Turillazzi S, Beani L. 2004. Social wasps desert the colony and aggregate outside if parasitized: parasite manipulation? Behavioral Ecology, 15, 1037–1043. [Google Scholar]
- Kanzaki N, Makino SI, Kosaka H, Sayama K, Hamaguchi K, Narayama S. 2023. Nematode and strepsipteran parasitism in bait-trapped and hand-collected hornets (Hymenoptera, Vespidae, Vespa). Insects, 14, 398. [Google Scholar]
- Kathirithamby J. 1989. Review of the order Strepsiptera. Systematic Entomology, 14, 41–92. [Google Scholar]
- Kathirithamby J. 2009. Host–parasitoid associations in Strepsiptera. Annual Review of Entomology, 54, 227–249. [CrossRef] [PubMed] [Google Scholar]
- Kathirithamby J. 2018. Biodiversity of Strepsiptera. Insect Biodiversity: Science and Society, 2, 673–703. [Google Scholar]
- Kim IK, Kim CJ, Choi JH, Kang HJ, Choi MB. 2025. Stylopization by Xenos spp. (Xenidae, Strepsiptera) in invasive alien hornet, Vespa velutina, in South Korea. Parasite, 32, 10. [Google Scholar]
- Kinzelbach R. 1971. Morphologische Befunde an Fächerflüglern und ihre phylogenetische Bedeutung (Insecta: Strepsiptera). Stuttgart: Schweizerbart’sche Verlagsbuchhandlung. [Google Scholar]
- Kristensen NP. 1991. Phylogeny of extant hexapods. In The Insects of Australia: a textbook for students and research workers (Vol. I, 2nd ed.). Melbourne: Melbourne University Press. pp. 125–140. [Google Scholar]
- Kudô K, Oyaizu W, Kusama R, Yamagishi K, Yamaguchi Y, Koji S. 2024. Mating and post-hibernation ovarian development in stylopized and non-stylopized queens of the hornet Vespa analis (Hymenoptera: Vespidae). Ethology Ecology & Evolution, 36, 86–95. [Google Scholar]
- Kudô K, Oyaizu W, Kusama R, Yamagishi K, Yamaguchi Y, Koji S. 2024. Effect of environmental factors on strepsipteran parasite abundance in vespine wasps (Hymenoptera: Vespidae). Sociobiology, 71, e10347. [Google Scholar]
- Lioy S, Laurino D, Capello M, Romano A, Manino A, Porporato M. 2020. Effectiveness and selectiveness of traps and baits for catching the invasive hornet Vespa velutina. Insects, 11, 706. [CrossRef] [PubMed] [Google Scholar]
- Makino S. 2001. Seasonal changes in levels of parasitism and sex ratio of Xenos moutoni du Buysson (Strepsiptera, Stylopidae) in the Japanese hornet, Vespa analis insularis Dalla Torre (Hymenoptera, Vespidae), collected with attractant traps. Tijdschrift voor Entomologie, 144, 217–222. [CrossRef] [Google Scholar]
- Makino S, Yamaura Y, Yamauchi H. 2010. Smaller nests of the hornet Vespa analis (Hymenoptera, Vespidae) are more severely affected by the strepsipteran parasite Xenos moutoni (Strepsiptera, Stylopidae) than are larger nests. Insectes Sociaux, 57, 83–90. [CrossRef] [Google Scholar]
- Makino SI. 2025. Levels of parasitism by Xenos oxyodontes (Strepsiptera, Stylopidae) among adult hornets and its voltinism in the host Vespa analis (Hymenoptera, Vespidae). Sociobiology, 72, e11306. [Google Scholar]
- Makino SI, Kawashima M, Kosaka H. 2011. First record of occurrence of Xenos moutoni (Strepsiptera; Stylopidae), an important parasite of hornets (Hymenoptera: Vespidae: Vespa), in Korea. Journal of Asia-Pacific Entomology, 14, 137–139. [Google Scholar]
- Makino SI, Yamashita Y. 1998. Levels of parasitism by Xenos moutoni du Buysson (Strepsiptera, Stylopidae) and their seasonal changes in hornets (Hymenoptera: Vespidae, Vespa) caught with bait traps. Entomological Science, 1, 537–543. [Google Scholar]
- Masciocchi M, Mattiacci A, Villacide JM, Buteler M, Porrino AP, Martínez AS. 2023. Sugar responsiveness could determine foraging patterns in yellowjackets. Scientific Reports, 13, 20448. [Google Scholar]
- Matsuura M, Yamane S. 1990. Biology of the vespine wasps. Berlin, Heidelberg: Springer-Verlag. [Google Scholar]
- Nain D, Rana A, Raychoudhury R, Sen R. 2024. Morphology, biology and phylogeny of Xenos gadagkari sp. nov. (Strepsiptera: Xenidae): an endoparasite of Polistes wattii (Hymenoptera: Vespidae). Zootaxa, 5493, 561–576. [Google Scholar]
- Nakase Y, Kato M. 2013. Cryptic diversity and host specificity in giant Xenos strepsipterans parasitic in large Vespa hornets. Zoological Science, 30, 331–336. [CrossRef] [PubMed] [Google Scholar]
- Oyaizu W, Kudô K. 2013. Seasonal changes in the number of vespine wasps and levels of parasitism by Xenos moutoni (Strepsiptera, Stylopidae) collected with attractant traps in Matsunoyama forest, Tokamachi city, Japan. Bulletin of the Faculty of Education (Natural Sciences), Niigata University, 6, 49–57 [in Japanese]. [Google Scholar]
- Pohl H, Beutel RG. 2008. The evolution of Strepsiptera (Hexapoda). Zoology, 111, 318–338. [CrossRef] [PubMed] [Google Scholar]
- Rossini L, Contarini M, Delfino I, Speranza S. 2025. Does insect trapping truly measure insect populations? Agricultural and Forest Entomology, 27, 329–339. [Google Scholar]
- Schrader SH. 1924. Reproduction of Acroschismus wheeleri Pierce. Journal of Morphology and Physiology, 39, 157–197. [Google Scholar]
- Tatsuta H, Makino SI. 2003. Rate of strepsipteran parasitization among overwintered females of the hornet Vespa analis (Hymenoptera: Vespidae). Environmental Entomology, 32, 175–179. [CrossRef] [Google Scholar]
Cite this article as: Kim J, Kim CJ, Choi MB & Kwon O. 2026. Phenology and parasitism rates of Xenos oxyodontes and Xenos moutoni (Strepsiptera: Xenidae) in Vespa (Hymenoptera: Vespidae) in South Korea. Parasite 33, 28. https://doi.org/10.1051/parasite/2026028.
All Tables
Diagnostic criteria for reproductive and developmental stage assignment in Xenos spp.
Number and stylopization status of Vespa individuals collected in traps across South Korea over fifteen years, organized by Vespa and Xenos species.
Colony composition and stylopization in 22 V. analis nests collected from Muui-do, Incheon, South Korea on September 2, 2020.
All Figures
|
Figure 1 Spatial distribution of 677 trapping sites in South Korea (2008–2023). Blue squares represent Malaise trap sites, sampled from 2008 to 2018, and red circles indicate sugar-attractant trap sites, sampled from 2019 to 2023. Shaded areas approximate the year-specific sugar attractant trap survey coverage over the 2019–2023 period, with the years specified on the map. |
| In the text | |
|
Figure 2 A female Xenos moutoni stylopizing a host hornet, Vespa crabro, protruding from the host abdomen. |
| In the text | |
|
Figure 3 Monthly life-stage compositions of female (A) Xenos moutoni and (B) Xenos oxyodontes individuals captured by traps in South Korea. The lines (top) indicate the monthly number of captured females (n), and the stacked bars (bottom) show the within-month proportional compositions of female reproductive stages. |
| In the text | |
|
Figure 4 Spatial distributions of the GLM-based, host-capture–adjusted expected stylopized-host counts ( |
| In the text | |
|
Figure 5 (A) Number of V. analis individuals stylopized by X. oxyodontes in each of the 22 colonies examined. Dark grey bars represent females, and white bars represent males. (B) Colony-level stylopization rate of V. analis by X. oxyodontes, calculated as the proportion of stylopized individuals among all adults examined in each nest. Each dot represents one V. analis colony, and the raincloud plot shows the distribution of colony-level stylopization rates. |
| In the text | |
|
Figure 6 Sex-specific developmental stage composition of X. oxyodontes in stylopized V. analis individuals collected from 22 nests. |
| In the text | |
|
Figure 7 Representative developmental stages of Xenos oxyodontes recovered from inside and outside Vespa analis nests. Vitellogenic females and not-emerged males were detected in both inside- and outside-nest samples, suggesting that nest departure may occur at or around these stages. Abbreviations: WB, whole body (female); Co, cephalothorax (female); HA, host abdomen; EP, empty puparium (male); P, puparium (male); WBP, whole body in puparium (male). |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.