| Issue |
Parasite
Volume 31, 2024
|
|
|---|---|---|
| Article Number | 72 | |
| Number of page(s) | 14 | |
| DOI | https://doi.org/10.1051/parasite/2024072 | |
| Published online | 19 November 2024 | |
Research Article
Culicoides and midge-associated arboviruses on cattle farms in Yunnan Province, China
Culicoïdes et leurs arbovirus associés dans les fermes d’élevage de bovins de la province du Yunnan, en Chine
1
Yunnan Tropical and Subtropical Animal Virus Diseases Laboratory, Yunnan Animal Science and Veterinary Institute, Fengyu Road, Panlong District, Kunming 650224, Yunnan, China
2
Key Laboratory of Transboundary Animal Diseases Prevention and Control (Co-construction by Ministry and Province), Ministry of Agriculture and Rural Affairs, Fengyu Road, Panlong District, Kunming 650224, Yunnan, China
3
Research Institute for the Environment and Livelihoods, Charles Darwin University, Ellengowan drive, Casuarina, NT 0810, Australia
4
Center for Animal Disease Control and Prevention, Xinxi Street, Lufeng 651200, Yunnan, China
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
(Ying-Liang Duan); This email address is being protected from spambots. You need JavaScript enabled to view it.
(Le Li)
Received:
9
July
2024
Accepted:
30
October
2024
Abstract
Culicoides spp. (Diptera: Ceratopogonidae) are small biting midges, some of which are the vectors of arboviruses affecting livestock, i.e., African horse sickness virus (AHSV), bluetongue virus (BTV), and epizootic hemorrhagic disease virus (EHDV). Yunnan Province, located in southwestern China, has a history of high prevalence of arboviruses. The diversity and abundance of Culicoides was observed between March 2022 and March 2023 on two cattle farms in Lufeng County of Yunnan Province, China and virus isolation and PCR detection were attempted from cattle blood and Culicoides spp. collected from the farms. Light trap collections contained 19 species of Culicoides belonging to 8 subgenera and one unplaced species group and were dominated by C. oxystoma (63.4%), C. imicola (16.2%), C. arakawae (13.4%), C. sp. near newsteadi (2.3%), and C. orientalis (1.7%). A total of 8,343 Culicoides were used for statistical analysis; from these collections 997 Culicoides specimens belonging to 10 species were screened for the presence of BTV, EHDV, Yunnan Orbivirus (YUOV), and Yongshan totivirus (YSToV) using reverse transcription quantitative PCR (RT-qPCR). One strain of YUOV was isolated from cattle blood and 7 isolates of YSToV were isolated from 5 different species of Culicoides. BTV and YSToV were detected from 2 and 3 pools of parous C. imicola specimens by RT-qPCR, respectively, which is the first report of a totivirus to be associated with Culicoides. Culicoides imicola is likely to be the major vector of Culicoides-borne arboviruses in Lufeng County, which is a relatively dry locality, and adult C. imicola may play a role of BTV overwintering.
Résumé
Les culicoïdes (Diptera : Ceratopogonidae) sont de petits moucherons piqueurs, dont certains sont les vecteurs d’arbovirus affectant le bétail, à savoir le virus de la peste équine africaine (AHSV), le virus de la fièvre catarrhale (BTV) et le virus de la maladie hémorragique épizootique (EHDV). La province du Yunnan, située dans le sud-ouest de la Chine, a un historique de prévalence élevée d’arbovirus. La diversité et l’abondance des Culicoides spp. ont été observées entre mars 2022 et mars 2023 dans deux fermes d’élevage de bovins du comté de Lufeng, dans la province du Yunnan, en Chine, et l’isolement du virus et la détection par PCR ont été tentés à partir du sang de bovins et des Culicoides spp. collectés dans les fermes. Les collections de pièges lumineux contenaient 19 espèces de Culicoides appartenant à 8 sous-genres et un groupe d’espèces non placé, et étaient dominées par C. oxystoma (63,4 %), C. imicola (16,2 %), C. arakawae (13,4 %), C. sp. proche de newsteadi (2,3 %) et C. orientalis (1,7 %). Un total de 8343 culicoïdes ont été utilisés pour l’analyse statistique; à partir de ces collections, 997 spécimens de Culicoides appartenant à 10 espèces ont été examinés pour la présence de BTV, EHDV, Yunnan Orbivirus (YUOV) et Yongshan Totivirus (YSToV) en utilisant la PCR quantitative à transcription inverse (RT-qPCR). Une souche de YUOV a été isolée du sang de bovin et 7 isolats de YSToV ont été isolés de 5 espèces différentes de Culicoides. Le BTV et le YSToV ont été détectés respectivement dans 2 et 3 pools de spécimens pares de C. imicola par RT-qPCR, ce qui constitue le premier signalement d’un totivirus associé à Culicoides. Culicoides imicola est probablement le principal vecteur d’arbovirus transmis par culicoïdes dans le comté de Lufeng, qui est un endroit relativement sec, et les adultes de C. imicola peuvent jouer un rôle dans l’hivernage du BTV.
Key words: Culicoides / bluetongue virus / Yunnan Orbivirus / Yongshan totivirus / Overwinter
Edited by: Jérôme Depaquit
Ying-Liang Duan and Zhan-Hong Li contributed equally to this work.
© Y.-L. Duan et al., published by EDP Sciences, 2024
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
Culicoides (Diptera: Ceratopogonidae) are small biting insects and at least 40 of the more than 1,300 species of Culicoides worldwide are reported as potential vectors for arboviruses [7, 39], including several important livestock viruses such as African horse sickness virus (AHSV), bluetongue virus (BTV), and epizootic hemorrhagic disease virus (EHDV) [36, 42, 49]. Recently, six species, i.e., Culicoides tainanus Kieffer, C. sp near obsoletus Meigen, C. imicola Kieffer, C. jacobsoni Macfie, C. orientalis Macfie, and C. oxystoma Kieffer, have been shown to carry arboviruses including BTV, Tibet orbivirus (TIBOV), EHDV, Banna virus (BAV), and Yunnan Orbivirus (YUOV) in Yunnan Province, China [11, 13, 16, 18, 24, 31].
Almost all members of the genus Orbivirus of Sedoreoviridae are arboviruses, for example AHSV, BTV, EHDV, Palyam virus, TIBOV, and YUOV [4, 26, 30, 49, 57, 58]. Some of these, like BTV and EHDV, mainly infect ruminants such as cattle, sheep, goats and deer, and are transmitted by Culicoides [12, 39, 49].
Totiviridae is composed of a group of viruses with a single dsRNA segment and parasitizing fungi, yeasts, and protozoa [26, 60]. A series of totivirus-like viruses, such as Yongshan totivirus (YSToV), Yuanmou totivirus (YMToV), Tianjin totivirus, and Shanghai totivirus have been reported in China over the past two decades [45, 46, 64]; however, these viruses came from mosquitoes instead of the known hosts of Totiviridae.
In subtropical and temperate zones, summer and autumn are the seasons when BTV is most prevalent among ruminants [35, 48, 63, 67]; however, the virus is often found to be absent during winter yet emerges again in the next warm season [35, 48]. The mechanism of BTV overwintering has been studied by several researchers, but has never been resolved [20, 41, 44, 48, 52, 53, 59].
Lufeng County of Yunnan Province, China is a relatively dry locality and experiences hot summers and cold winters. Quite a few bovine and goats positive for BTV antibodies were reported in three counties (Chuxiong County, Shuangbai County, and Yuanmou County) adjacent to Lufeng County in 2002 [8]. But so far, no disease of bovines, goats or sheep has been publicly reported from Lufeng County. However, five cattle from a farm in Lufeng County had died of disease in the summer in 2021. Subsequently, blood samples of 24 subclinical and healthy cattle were collected in 2021 and submitted to the Yunnan Tropical and Subtropical Animal Virus Diseases Laboratory. Approximately 5 of 24 were positive for BTV RNA, and 4 of 24 were positive for EHDV RNA, through RT-qPCR detection by Le Li et al. (unpublished). This event prompted us to investigate the Culicoides and the arboviruses, especially BTV and EHDV, on this farm in the next year. Moreover, this is an ideal locality to investigate the overwintering situation of arboviruses in Lufeng County.
This study reports the species diversity and abundance of Culicoides and virus isolation and detection from cattle and Culicoides on two cattle farms in Lufeng County, Yunnan Province, China.
Materials and methods
Ethics approval and consent to participate
The procedure for sampling cattle blood was approved by the Ethics Committee of the Yunnan Animal Science and Veterinary Institute under number YNASVI01-2022LL.
Collecting samples
Two cattle farms (farms A and B) in Caiyun town, Lufeng County, Yunnan Province, China were set up as collection sites (Fig. S1A). There is a 3.5 km straight line distance between the two farms. Farms A and B fed approximately 380 cattle in three adjacent pens and 200 cattle in two adjacent pens, respectively (Table S1). As mentioned above, five cattle on farm A had died of disease in 2021. Thereafter, hygiene practices on both farms included twice weekly spraying of pesticides in the pens and regular removal of dung. On each farm, two battery powered UV-light traps (Yaoyu electronics Co. Ltd., Zhangzhou, China) were set; one trap captured living midges into a dry bottle with fresh leaves to provide humidity, while the other collected midges into a bottle containing 75% ethanol (Table S2). The traps ran from approximately 4:00 pm (before sunset) to 9:00 am the following day. Living midges were processed for virus isolation soon after collection, while the midges in ethanol were stored at 4 °C until processed. Culicoides were collected on one night each month between March 2022 and March 2023 (Table S2).
Additionally, 39 EDTA+ blood samples were collected from 20 cattle on Farm A on 3 occasions between March 4 and June 1, 2022 (Table S2, Fig. S1B), and stored at 4 °C.
Sorting of Culicoides
Culicoides collected in ethanol were identified according to the morphological keys of Wirth [61] and Yu et al. [66], and graded into blood fed, parous + gravid and nulliparous females, and males. Parous status was identified by the presence of burgundy pigment in the abdomen [19]. Representative specimens of all species were mounted onto glass slides following the methods described by Bellis et al. [6]. All specimens collected were counted and the male rate (= male midges/total midges), blood fed rate (= blood fed midges/female midges), and nulliparous rate [= nulliparous midge/(parous midges + nulliparous midges)] were calculated for each collection.
Weather data
Daily weather data for Lufeng County were collected from the website of Baidu weather [2]. Daily precipitation data in Lufeng County were not available, so precipitation was estimated by the categories of weather. Rainfall over each 24-hour period was classified into five categories: rainstorm (50.0–99.9 mm), heavy rain (25.0–49.9 mm), moderate rain (10.0–24.9 mm), and light rain (0.1–9.9 mm) or overcast by the China Meteorological Administration [1]. No rainstorm or heavy rain was reported in this county during the period we studied. To estimate daily precipitation quantitatively, rainfall scores were designed as 4 points for moderate rain, 2 points for light rain, 0.5 points for overcast or fog, and 0 point for sunny or cloudy.
Sample preparation for viral isolation
Prior to October 2022, live midges were prepared for viral isolation on the day of collection but thereafter (between October 2022 and March 2023) they were incubated at no less than 12 °C for 24 h before processing.
Midges were immobilized at −20 °C for at least 20 min and parous + gravid female Culicoides without a visible blood meal were selected for viral isolation. Conspecific specimens were pooled and placed into 1.5 mL EP tubes with 1 mL sterilized phosphate buffered saline (PBS). Each pool contained between 1 and 35 specimens. PBS was removed following soft shock using a vortex machine for 30 s and centrifugation (1,850 × g, 2 min), then the operation was repeated. Subsequently, 600 μL minimum essential medium (MEM) and two sterile steel balls (Φ3 mm) were added to each tube. Specimens were homogenized at 25 Hz for 25 min using a multiple functional homogenizer TissueLyser II (QIAGEN, Hilden, Germany). The homogenates were centrifuged (1,850 × g, 2 min), and the supernatants were used for inoculation.
For blood samples, 1 mL blood was centrifuged at 300 × g for 5 min, the plasma was removed and the red blood cells were suspended in 1 mL sterile PBS. After centrifugation, PBS was replaced with 1 mL sterile PBS and then the operation was repeated, after which 1 mL of sterile double distilled water was added to suspend the cell pellet, and the suspension was placed on ice for 10 min prior to inoculation onto cell lines.
Viral isolation
Baby hamster kidney (BHK-21) and Aedes albopictus mosquito (C6/36) cell lines were cultured in MEM with 5% fetal bovine serum (FBS), 100 U/mL penicillin and 100 μg/mL streptomycin (Gibco, Thermo Fisher Scientific, Grand Island, NY, USA) at 37 °C and 28 °C, respectively. For viral isolation, BHK-21 and C6/36 cells were prepared individually in 6-well plates and 300 μL of supernatant prepared from midges or blood described above were added to each well. The medium was replaced by 3 mL MEM with 1% FBS at 2 h post inoculation. Infected cells were cultured for 7 days and examined daily for obvious cytopathic effect (CPE). Cells exhibiting CPE were removed to a 5 mL tube and stored at −80 °C. If after 7 days no CPE appeared, the entire plate was frozen and thawed twice, centrifuged at 1,850 × g for 3 min and 300 μL of supernatant were inoculated onto new cells and cultured for a further 7 days. Cells were passaged in this manner 3 times then discarded.
Extraction of nucleic acids from viral isolates
All cell cultures exhibiting CPE were subjected to viral nucleic acid extraction. Briefly, nucleic acids were extracted from 200 μL of cellular medium using a MiniBEST Viral RNA-DNA Extraction Kit (#2158, Takara, Dalian, China), according to the manufacturer’s instructions. Nucleic acids were eluted by 35 μL/sample RNase-free water and kept at −80 °C until use. The samples from the positive cell cultures were used for metagenomic sequencing and viral identification by PCR, respectively.
Metagenomic sequencing
An aliquot of 10 μL of nucleic acid from up to 8 isolates was pooled and sent to MAGIGENE Science and Technology Ltd for sequencing. Briefly, a DNA library was constructed using an NEB Next® Ultra II™ DNA Library Prep Kit for Illumina (#E7645S, New England Biolabs, Ipswich, MA, USA), according to the manufacturer’s instructions; the quality of the library was checked by Agilent 4200 TapeStation (Agilent Technologies, Santa Clara, CA, USA) using a Qubit® dsDNA HS Assay Kit (Life Technologies, Carlsbad, CA, USA). Samples were sequenced by a Novaseq 6000 System (Illumina, San Diego, CA, USA) using 150 bp paired-end pattern.
Phylogenetic analysis
Two contigs acquired from metagenomic sequencing, namely contig-214 for the YUOV T2 (VP2) gene and contig-S5 for the YSToV partial genome (Table S3), were used for phylogenetic analysis. Reference sequences of similar viruses were downloaded from NCBI. Sequences were aligned by MUSCLE (Codons) and phylogenetic trees were constructed using a Neighbor-Joining (NJ) algorithm (bootstrap = 1,000), using MEGA 11 software [23]. As some sequences were not complete, the protruding ends of all sequences were truncated following alignment so that they were all of equal length. For T2 gene analysis, the sequences were truncated to begin at the 4th codon and end at the penultimate codon with a length of 2,811 bp for YUOV and middle point orbivirus (MPOV) and 2,694 bp for BTV. For RNA dependent RNA polymerase (RdRP) analysis, sequences were truncated to start at the 1st codon and end at the penultimate codon according to YSToV strain Yunnan/2018, and with a length of 2,031 bp.
Extracting nucleic acid from midges
Conspecific pools of no more than 5 blood fed female or no more than 10 parous + gravid female midges were tested for the presence of viruses. Each pool of specimens was digested non-destructively in 55 μL tissue lysis buffer (TIANGEN, Tiangen, Beijing, China) containing 0.2 mg/mL proteinase K (TIANGEN) at 25 °C for 16 h [15]. A MagMAX™ Express-24 machine (Ambion, Thermo Fisher Scientific, Waltham, MA, USA) coupled with KingFisher plate (Thermo Fisher Scientific) and MagMAX™-96 Viral RNA Isolation kit (Ambion) were used to extract the nucleic acid from the supernatant, according to the manufacturer’s instructions, and the nucleic acids were eluted in 50 μL/well elution buffer. For preliminary screening, a 10 μL aliquot from 4 pools of insects were pooled and tested (Fig. S1C) and positive pools were investigated further by testing 40 μL of lysate of each of the 4 single pools comprising the positive group (Fig. S1D).
Reverse transcription and polymerase chain reaction (RT-PCR)
The cDNA of all viral isolates was synthetized by 10 μL of RNA by ProFlex PCR System (ABI, Foster City, CA, USA) using a PrimeScrip 1st Strand cDNA Synthesis Kit and random primers (#6110A, Takara), according to the manufacturer’s instructions.
To identify YUOV and YSToV, 2.5 μL of viral cDNA was added to 22.5 μL of PCR reaction system confected using PrimeSTAR HS (#R040Q, Takara) and primers designed in this study (Table S4), according to the manufacturer’s instructions. The PCR cycling program consisted of: denaturation at 95 °C, 1 min, then denaturation at 95 °C for 10 s, annealing at 55 °C for 5 s, extension at 68 °C for 80 s (YUOV) or 1 min (YSToV) for 30 cycles; and a final extension at 68 °C for 1 min, with storage at 4 °C.
PCR amplified DNA fragments (5 μL per sample) were separated by electrophoresis in 1.5% agarose gel with dye GoldView-II, and screened by a Gel Doc XR+ System (Bio-Rad, Hercules, CA, USA).
Detecting virus in midges by RT-qPCR
A one-step RT-qPCR method was used to scan for 4 viruses (BTV, EHDV, YUOV, and YSToV) in midges. The primers and probes for BTV [25] and EHDV [65] were used from previous reports, while the primers and probes for YUOV and YSToV were novel (Table S4). A 2 μL aliquot of midge sample was added to 20 μL of reaction solution prepared using a Quant One Step PrimeScript RT-PCR Kit (Takara), according to the manufacturer’s instructions. The RT-qPCR was performed on a Fast7500 Realtime PCR machine (ABI) at the following cycling conditions: 42 °C, 5 min; 95 °C, 10 s; 95 °C for 10 s, 60 °C for 34 s, 40 cycles. Fluorescence was measured at the end of each extension step.
For the primary scan, samples were detected by dual channel RT-qPCR for BTV/EHDV and YUOV/YSToV, respectively (Fig. S1C). For further scanning, a candidate sample from a single pool was detected by RT-qPCR for single target in triplicate, according to the result of primary RT-qPCR (Fig. S1D).
Confirming the BTV in Culicoides
To confirm the positive results of RT-qPCR experiments for BTV, primers against BTV NS3 were designed based on the sequence of strain BTV4/YTS-4 (JX560422.1) (Table S4). Detailed methods and results are described in Supplemental File 1.
Results
Culicoides diversity and abundance
Collections yielded 19 species of Culicoides belonging to 8 subgenera (Avaritia, Beltranmyia, Culicoides, Hoffmania, Meijerehelea, Monoculicoides, Oecacta, and Remmia) and one species group (Clavipalpis group) (Table 1, Fig. 1). A total of 8,343 specimens were collected with UV traps with ethanol and used for statistical analysis, and 754 parous/gravid female specimens without blood meal from living collections were used for viral isolation. Specimens of C. sp near obsoletus, a species previously reported in Yunnan [13], were only found in the live collections for viral isolation. A novel Culicoides species belonging to subgenus Culicoides and with similar morphology to C. neilamensis Liu & Deng [32] was found. Seasonal abundance of individual species of Culicoides on the two farms was in general agreement with summer peaks evident in all species (Fig. 2). According to the number of specimens collected on both farms over the year, the dominant species was C. oxystoma (63.4%), followed by C. imicola (16.2%), C. arakawae Arakawa (13.4%), C. sp near newsteadi Austen (2.3%; a species reported as C. punctatus Latreille by us before) and C. orientalis (1.7%), with the remaining species contributing less than 1% (Table 1). Midge abundance was generally quite low with most collections on both farms containing no midges, and all but 5 containing less than 300 specimens (Fig. 2).
 |
Figure 1 Wings of represented female specimens. The wings of represented Culicoides species collected from Lufeng County in this study, scale bar = 0.5 mm. |
 |
Figure 2 Abundance of Culicoides species on two farms in Lufeng Province, Yunnan, China. Each cell represents a collection and the color reflects the Culicoides abundance in each collection. |
Diversity and abundance of Culicoides collected over a 12-month period from 2 farms in Lufeng County, Yunnan Province, China.
Annual weather data between March 1, 2022 and March 31, 2023 in Lufeng showed that daily maximum temperatures ranged from 6 °C to 32 °C, and daily minimum temperatures ranged from 0 °C to 25 °C, while the daily temperature difference ranged from 1 °C to 23 °C. Minimum temperatures changed steadily with the season and rainfall usually lowered the maximum temperature and decreased the average daily difference in temperature (Fig. 3C).
 |
Figure 3 Seasonal abundance of Culicoides and weather on 2 farms in Lufeng County, Yunnan, China. (A) The relative proportion of female midges belonging to the major species present in collections from Mar 2022 to Mar 2023. (B) Total midge abundance (log10 (n + 1)) on Farm A and Farm B, and the average abundance at the two farms from Mar 2022 to Mar 2023. (C) Daily maximum and minimum temperature (TEM) and rain score between March 1, 2022 and March 31, 2023 in Lufeng County. |
Generally, all species of Culicoides were active between late spring and autumn (between April and October) peaking in the summer (Figs. 2 and 3B), showing a correlation with both temperature and rainfall (Figs. 3B and 3C). Culicoides oxystoma and C. arakawae both showed peak abundance in summer, while C. imicola was found in every month, except January 18; C. sp. near newsteadi became the dominant species in winter (Fig. 3A).
Proportions of male, blood fed and nulliparous Culicoides
Male specimens for 11 species were collected (Table 2). Culicoides circumscriptus Kieffer had the highest male: female ratio but only 2 specimens were collected; C. homotomus Kieffer and C. oxystoma had average male: female ratios above 30%, while the remainder had ratios below 20%.
Proportion of male specimens in UV trap collections of Culicoides collected on 2 farms over 12 months in Lufeng County, Yunnan Province, China.
Seasonal changes in the blood fed and nulliparous rate of C. arakawae, C. imicola, C. oxystoma, and total Culicoides between April 28 and December 15 were studied (Fig. 4) as too few specimens of other species were collected to analyze. The blood fed rate of all species ranged from 5.1% to 14.1% between April 28 and November16, while no blood fed Culicoides were found in December (Fig. 4A). The proportion of blood fed specimens of C. arakawae was lower than that of other species, and was zero in most cases, while the proportion of blood fed C. imicola was higher than other species, often around 30% (Fig. 4A). The nulliparous rates of Culicoides were not significantly correlated with the season; generally, the nulliparous rate of C. arakawae was lower than that of other species, while the values of C. imicola and C. oxystoma were synchronous with the total Culicoides (Fig. 4B).
 |
Figure 4 Proportions of blood fed and nulliparous females. The average percentages (mean ± SD) of blood feeding (A) and nulliparous female (B) Culicoides collected on 2 farms in Lufeng county, Yunnan, China between April and December 2022. |
Viral isolates
Virus isolation was attempted on 39 blood samples from 20 cattle and 754 specimens of Culicoides in 95 pools (Table S5), and in total, 8 viruses were isolated. These isolates were acquired from C6/36 culture and none produced CPE in BHK-21 cells. Metagenomic sequencing produced 11 confirmed contigs from YUOV and YSToV (Table S3), which cued that the unknown viral isolates belonged to these two viruses. Subsequently, the isolates were identified by specific RT-PCR (Fig. S2) and confirmed by sequencing using PCR amplified fragments (Table S3). As a result, a single isolate of YUOV was obtained from cattle and 7 isolates of YSToV were isolated from 7 pools of conspecific midges representing 5 species, i.e., C. arakawae, C. asiana Bellis, C. orientalis, C. oxystoma, and C. tainanus (Table 3). The 7 YSToV isolates belonged to a single strain based on the sequences of PCR fragments.
Viruses isolated from pools of Culicoides and cattle blood collected from 2 farms in Lufeng County, Yunnan Province, China.
In the phylogenetic analysis, the YUOV isolate (contig-214) grouped with a cluster of other YUOV and MPOV viruses (Fig. 5A). The YSToV LF2022 (contig-S5) grouped closely with one other strain of YSToV but also with strains of Yuanmou totivirus (YMToV), Tianjin totivirus (TJToV), Shanghai totivirus (SHToV), and 3 strains of Omono River virus (OMRV) (Fig. 5B).
 |
Figure 5 Phylogenetic analysis for YUOV and YSToV, and related viruses. Phylogenetic trees of T2 genes of YUOV, MPOV, and BTV (A), and RdRP genes of YSToV and close totivirus (B) were constructed by NJ algorithm. The sequences of our isolates (contig-214, contig-S5) were marked by circles. Bootstrap values less than 70% were omitted. |
RT-qPCR scanning for viruses in midges
A total of 997 specimens placed into 144 pools of conspecific midges representing 10 species were tested by RT-qPCR for the presence of BTV, EHDV, YSToV, and YUOV (Table S6). Two pools of parous C. imicola were positive for BTV although the Ct values were high (31.5 and 36.4, respectively) while 3 pools of parous C. imicola were positive for YSToV (Table 4). The positive results of BTV in C. imicola were confirmed by amplification and sequencing of the BTV NS3 fragments (Supplemental File 1). The midges collected on November 16 had obviously lower BTV abundance (Table 4), and the positive rate of BTV among C. imicola was approximately 0.6% (2/331) (Table S6).
RT-qPCR results from pools of parous + gravid Culicoides collected in Lufeng County, Yunnan Province, China.
Discussion
Lufeng County, located in the central part of Yunnan, experiences a relatively dry climate with warm days (in summer and autumn) and cold nights. The Ailao Mountain range prevents the northward movement of the summer monsoon from the Indian Ocean, so north of this range, the climate is relatively dry. However, cattle dying of disease on a farm (Farm A) in Lufeng County in 2021 prompted us to begin an investigation of Culicoides and midge-associated arboviruses in the following year.
More than 9,000 specimens of Culicoides representing 19 species were collected from two cattle farms over a 12-month period. It is possible that the hot, dry days in summer and autumn, coupled with cold nights during winter, and regular spraying of pesticides on dung may have contributed to the low abundance of midges on the two farms. The dominance of C. oxystoma, C. imicola, and C. arakawae has also been observed on livestock farms in some parts of southern Asia [3, 10, 14, 16, 22]. However, this county has distinctive diversity of Culicoides compared to other counties in Yunnan.
Notably absent from collections in this study were species belonging to C. subgenus Trithecoides, as these species are common and often dominant on livestock farms in south Yunnan and other places in southern Asia [16, 22, 61]. Similarly, only 5 specimens of subgenus Hoffmania were collected; however, this is consistent with previous reports from Yunnan that C. (Hoffmania) species prefer altitudes below 11,00 m [15], while these collection sites are approximately 1,400 m above sea level. Previous studies have reported that C. tainanus is widespread and common in Yunnan, while C. imicola is relatively rare [13, 14, 16, 33]. At Lufeng, however, C. imicola are obviously more abundant than C. tainanus, although the reasons for this are unclear. This study provides only the second record of C. morisitai Tokunaga from Yunnan [33, 66]. It is interesting that C. morisitai was absent in summer and autumn in this investigation, while the researchers always collected Culicoides in these seasons, so this species might be missed.
Low numbers of specimens of a species close to C. obsoletus were collected but only in the live collections. DNA barcodes of this species differ significantly from European populations of C. obsoletus [13], suggesting that these represent two distinct species, despite their morphological similarity. Culicoides obsoletus has been reported from various places in Yunnan [10, 13, 33]. However, these reports are all based on morphological identification and need to be checked to ensure they do not belong to this cryptic species. Female specimens of several species in the Obsoletus complex are morphologically indistinguishable [38] so examination of male specimens will be needed to clarify the status of these species.
A single specimen found in this study appears to belong to an undescribed species belonging to C. subg. Culicoides. This new species belongs to a complex of species similar to C. nielamensis that we have reported from Shangri-La [13] and found at other high altitude sites in Yunnan. None of these species have the poststigmatic pale spot in cell r3 reaching vein M1 which is apparently a characteristic of C. nielamensis (Fig. 1) [13, 32]. Further study, including examination of reference specimens of C. nielamensis is needed to clarify the status of these species.
Culicoides punctatus and C. newsteadi identified in Europe were similar in morphology [5], and a similar species complex was reported in Asia [14, 27, 28]. A re-examination of the specimens previously reported as C. punctatus by Matsumoto et al. [37], Kim et al. [27, 28], and Duan et al. [14] and probably other studies in Asia, has prompted us to place these specimens closer to C. newsteadi than to C. punctatus in morphology (Fig. 1), although DNA barcode data indicate that Asian populations are actually different to both of these species [37] (Duan et al. unpublished data), so we prefer to refer to this species as C. sp. near newsteadi.
Three proven, C. actoni, C. fulvus and C. imicola [36, 47, 51], and several suspected vectors of BTV, which were shown to carry BTV in the field, C. circumscriptus, C. jacobsoni, C. sp near newsteadi, C. oxystoma, C. sp near obsoletus, and C. tainanus [13, 16, 21, 31] were collected at Lufeng, although the relative importance of the species in BTV epidemiology is difficult to assess. Only C. imicola was found to be carrying BTV in this study. Additionally, only low numbers of C. actoni, C. jacobsoni, C. sp. near newsteadi, C. sp near obsoletus, and C. tainanus and none of C. fulvus were tested so it is difficult to assess these species.
Compared to some studies, the Ct values (31.5 and 36.4) for BTV were relatively high in this study, which might be caused by a less optimal method of collecting midges, since systemic errors could be caused by the quality of specimen preservation, efficiency of nucleic acid extraction, or reagents for molecular experiment. In this study, midges for sorting were directly collected in a bottle loading 75% ethanol and connecting to the UV-trap. However, the ethanol began to volatilize as soon as the trap was set. So the trapped midges would soak in low concentrations of ethanol for hours before collecting. Virions in midges might be digested by intracellular proteinases and nucleases partly during the process of apoptosis or necrosis of midge cells. Therefore, the Ct values for BTV in Culicoides specimens might be underestimated in this study. Low Ct values (between 20 and 25) for BTV could be acquired in the experiments of artificial infection on Culicoides in laboratory [54, 56]. For field midges, collecting midges using a mesh bag under the UV-trap might be better to preserve the midges and viruses [13, 16]. While high Ct values (between 30 and 50) for BTV were acquired from some investigation for field collected midges [21, 31].
The life cycle of Culicoides undergoes 4 stages, namely egg, larva, pupa, and adult [48]. Adult female midges typically mate, take a blood meal, lay eggs, and then take another blood meal and produce eggs in a continuous cycle. Meantime, sucking blood is the critical stage to spread arbovirus among hosts of Culicoides. Therefore, blood feeding is the index of both midge reproduction and viral transmission, while nulliparous midges represent newborn adults. As male midges never suck blood, they will in theory appear near their habitat or plant food. The relatively high proportion of male specimens of C. imicola and C. tainanus was unexpected as they are usually uncommon in most collections in Yunnan [13, 16] and in South Korea [28].
Culicoides arakawae is considered to be a bird feeder [29]. Relatively high male: female ratios of this species compared to other species have been reported in light traps [28], suggesting either a greater attraction of males of this species to lights or cattle compared to males of other species. The very low number of blood fed female C. arakawae collected in Lufeng is likely due to their preference to feed on birds [29], although the possibility that blood fed females are not attracted to lights cannot be ruled out. The high abundance of this species is consistent with other studies [16, 28] and we assume this species is highly attracted to lights, even if traps are not set close to hosts. In this study, C. imicola had a higher proportion of blood fed females than other species, and although this does not necessarily equate to a high attack rate on cattle, it does suggest that this species is feeding on the cattle at this site and, given its proven status as a vector of BTV, likely involved in the transmission of virus.
BTV and EHDV are important livestock arboviruses [35, 43, 49, 62] that are prevalent among domestic ruminants in Yunnan Province but have not been reported to cause clinical disease [12, 17, 31] since the first bluetongue cases of 430 sheep in mainland China (Shizong County of Yunnan Province) in 1979 [67]. In summer 2021, 5 cattle died of disease on Farm A in Lufeng County of Yunnan, China. Subsequently, a few blood samples collected from the cattle on the same farm were positive for BTV, EHDV, or for both viruses (Li et al., unpublished). Therefore, the farm owner enhanced cleaning on the two farms (sites A and B). In this study, two pools of C. imicola from June 1, 2022 and November 16, 2022, respectively were positive for BTV. The absence of BTV in cattle blood and in blood fed midges suggests a low prevalence of BTV on these farms between March 2022 and March 2023. It seems that hygiene practices effectively reduced the prevalence of BTV and EHDV on this farm. The higher Ct value in the positive midge collected in November is consistent with the theory that the RNA polymerase of BTV would be inhibited below 10 °C [55].
BTV has to subsist on its hosts of ruminants and Culicoides. However, ruminants such as cattle will eliminate the invaded BTV by specific immunity within months since first infection [50], or killed by the disease in a short time. The Culicoides are short-lived, whether the BTV persistently infects single Culicoides or not. Therefore, BTV should continuously change host between ruminant and Culicoides. In subtropical and temperate zones, BTV prevalence among ruminants in summer and autumn is synchronized with the activity of Culicoides [35, 48]. However, cases of BTV decline as the abundance of Culicoides declines in winter. So far, the mechanism of BTV overwintering in temperate climates is uncertain [20, 48, 53]. As BTV does not have a DNA genome, it should not have latent infection patterns in ruminants, theoretically and empirically. But it was reported that BTV can be alive in sheep skin for 9 weeks, which is longer than the time of viremia [53]. There is no evidence of vertical transmission of BTV in Culicoides [44]. But BTV might overwinter through adult Culicoides [20, 52]. In this study, low levels of BTV RNA were detected from a pool of C. imicola in November, which partly supports the mechanism of BTV overwintering through adult Culicoides. Low levels of BTV in adult Culicoides are a balance of coexistence between BTV and Culicoides. Because lower abundance of virions means less harm for the host, this theoretically delayed the death of Culicoides caused by infection. Furthermore, adult Culicoides live longer at lower temperatures [34]. Although this investigation suggested that C. sp near newsteadi could endure lower temperatures compared with other species, according to increasing ratios of C. sp near newsteadi in winter; no BTV was detected in specimens from this species, which might be caused by a relatively lower amount.
YUOV was first identified in Yunnan, China [4] and, based on limited field research, was considered to be a mosquito-borne arbovirus [4, 40, 57]. The negative YUOV isolation and PCR results from Culicoides coupled with repeated isolation of virus from cattle blood suggest that cattle are a vertebrate host, but Culicoides are not important vectors of this virus. The phylogenetic analysis suggested that YUOV and MPOV found in Australia [9] were the same virus species.
The isolation of YSToV from 5 species of Culicoides was surprising as Totiviruses have so far only been associated with fungi, yeasts, and protozoa [26, 45, 46], so the presence of a totivirus in insects was surprising. It is possible that the YSToV isolates came from fungi present either internally or externally on the midges as reported for bats by Yang et al. [64]. However, YSToV was isolated in C6/36 mosquito cells in this study, which illustrates that YSToV can proliferate in insect cells. Additionally, some totiviruses have been isolated and detected frequently in mosquitos, suggesting more than a passing association with these insects [45, 46]. No vertebrate hosts have been found for these viruses so it is not clear if mosquitoes or midges are acting as vectors or are merely hosts of YSToV. The phylogenetic analysis suggested that YSToV was very closely related to some but not all strains of YMToV, TJToV, SHToV, and OMRV, suggesting that the relationships between these viruses are more complex than currently thought, and more work is required to clarify the status of these viruses.
All the YSToV isolates were acquired from the midges collected on July 6, 2022, suggesting a very high circulation of YSToV among midges in July. The cause is uncertain according to existing information. As the C. arakawae reached a very high proportion (>50%) on July 6, 2022 (Fig. 3A), C. arakawae might be a major host of YSToV, therefore increasing the circulation of YSToV among Culicoides in July. It is unknown whether fungi, yeasts, or protozoa are hosts of YSToV. Presumably high temperature and humidity would promote the growth of these hosts and increase the amount of virus in the environment and expose more midges to infection. However, the air humidity at the collection sites changes significantly during the day, so it is not possible to measure and compare the humidity changes month by month.
Conclusions
In total, 19 species of Culicoides belonging to 8 subgenera and one species group were found in Lufeng County, Yunnan, China between March 2022 and March 2023. The dominant species was C. oxystoma, followed by C. imicola, C. arakawae, C. sp near newsteadi, and C. orientalis. Three well-known BTV vectors (C. actoni, C. fulvus, and C. imicola) and 6 potential BTV vectors (C. circumscriptus, C. jacobsoni, C. sp near newsteadi, C. oxystoma, C. tainanus, and the C. sp near obsoletus) were collected. However, C. imicola was considered the major vector of Culicoides-borne arboviruses in this county, which is a relatively dry area with hot summers and cold winters. Furthermore, adult C. imicola may play an important role in BTV overwintering here. Culicoides arakawae is considered not to be a cattle feeder, although they can sometimes be abundant on cattle farms. One strain YSToV containing 7 viral isolates was the first totivirus to be isolated from Culicoides, but more work is required to clarify the status of these viruses, their hosts, and their ability to cause disease.
Acknowledgments
We thank L.X. Ma (the director of the two cattle farms) for permission to access his farms and sample his cattle.
Funding
This study was supported by the National Natural Science Foundation of China (32160846), and the Foreign Experts Project of Yunnan Province (202305AO350024).
Conflicts of interest
The authors declare that they have no conflict of interest.
Data availability statement
Data supporting the conclusions of this article are included within the article and its additional file. The datasets used and/or analyzed during the present study are available from the corresponding author upon reasonable request.
Author contribution statement
Conceptualization: Y.D. and L.L.; resource: all authors; investigation: Y.D. and Z.L.; formal analysis: Y.D. and Z.L.; visualization: Y.D.; writing – original draft: Y.D. and G.A.B.; writing – review & editing: Y.D. and G.A.B.; data curation: Y.D. and L.L.; funding acquisition: Y.D. All authors reviewed and approved the final manuscript.
Supplementary material
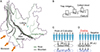 |
Figure S1: Locality map of Yunnan and flow chart of experiment. (A) Sketch map of Yunnan Province, China. Major rivers and mountains are shown roughly by green and grey lines, respectively. (B) Collecting samples on Farm A. (C) Preliminary RT-qPCR test for Culicoides. (D) Further RT-qPCR test for positive samples. |
 |
Figure S2: Identification of viral isolates by RT-PCR. |
Table S1: Details of collection sites in Lufeng County, Yunnan Province, China. Access Supplementary Material
Table S2: Blood samples and insect collections collected in this study. Access Supplementary Material
Table S3: High quality contigs and PCR fragments acquired in this study. Access Supplementary Material
Table S4: Novel primers and probes designed in this study. Access Supplementary Material
Table S5: Details of the pools of Culicoides used in virus isolation attempts. Access Supplementary Material
Table S6: Species composition and size of pools of Culicoides processed for the presence of virus by RT-qPCR. Access Supplementary Material
Supplemental File 1: Methods and results to confirm BTV in Culicoides. Access Supplementary Material
References
- Anonymous. 2022. Meteorological knowledge. Available from: http://www.qxkp.net/qxbk/qxsy/ [cited 5 Apr 2022]. [Google Scholar]
- Anonymous. 2023. Weathernews (Lufeng). Available from: https://weathernew.pae.baidu.com/weathernew/pc?query=%E6%A5%9A%E9%9B%84%E7%A6%84%E4%B8%B0%E5%A4%A9%E6%B0%94&srcid=4982&forecast=long_day_forecast [cited 31 Mar 2023]. [Google Scholar]
- Archana M, D’Souza PE, Renuka Prasad C, Byregowda SM. 2016. Prevalence of different species of Culicoides in Bangalore rural and urban districts of South India. Journal of Parasitic Diseases 40(3), 591–604. [CrossRef] [PubMed] [Google Scholar]
- Attoui H, Jaafar FM, Belhouchet M, Aldrovandi N, Tao S, Chen B, Liang G, Tesh RB, de Micco P, de Lamballerie X. 2005. Yunnan Orbivirus, a new Orbivirus species isolated from Culex tritaeniorhynchus mosquitoes in China. Journal of General Virology 86(Pt12), 3409–3417. [CrossRef] [PubMed] [Google Scholar]
- Augot D, Mathieu B, Hadj-Henni L, Barriel V, Zapata Mena S, Smolis S, Slama D, Randrianambinintsoa FJ, Trueba G, Kaltenbach M, Rahola N, Depaquit J. 2017. Molecular phylogeny of 42 species of Culicoides (Diptera, Ceratopogonidae) from three continents. Parasite 24, 23. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Bellis GA, Dyce AL, Gopurenko D, Mitchell A .2013. Revision of the immaculatus group of Culicoides Latreille (Diptera: Ceratopogonidae) from the Australasian Region with descriptions of two new species. Zootaxa 3680(1), 15–37. [CrossRef] [Google Scholar]
- Borkent A, Dominiak P. 2020. Catalog of the biting midges of the world (Diptera: Ceratopogonidae). Zootaxa 4787, 1–377. [CrossRef] [Google Scholar]
- Cao XP, Li CH, Li XL. 2002. Serologic investigation of bluetongue in bovine and goats/sheep in Chuxiong Prefecture of Yunnan Province. China Animal Health Inspection 19(9), 40. [Google Scholar]
- Cowled C, Melville L, Weir R, Walsh S, Hyatt A, Van Driel R, Davis S, Gubala A, Boyle D. 2007. Genetic and epidemiological characterization of Middle Point orbivirus, a novel virus isolated from sentinel cattle in northern Australia, Journal of General Virology 88(Pt 12), 3413–3422. [CrossRef] [PubMed] [Google Scholar]
- Di D, Li C, Li Z, Wang X, Xia Q, Mona S, Li B, Liu K, Shao D, Qiu Y, Soe-soe W, Yang S, Wei J, Ma Z. 2021. Detection of arboviruses in Culicoides (Diptera: Ceratopogonidae) collected from animal farms in the border areas of Yunnan Province, China. Journal of Integrative Agriculture 20(9), 2491–2501. [CrossRef] [Google Scholar]
- Duan Y, Yang Z, Bellis G, Xie J, Li L. 2022. Full genome sequencing of three Sedoreoviridae viruses isolated from Culicoides spp. (Diptera, Ceratopogonidae) in China. Viruses 14(5), 971. [CrossRef] [PubMed] [Google Scholar]
- Duan Y, Yang Z, Zhu P, Xiao L, Li Z, Li Z, Li L, Zhu J. 2022. A serologic investigation of epizootic hemorrhagic disease virus in China between 2014 and 2019. Virologica Sinica 37(4), 513–520. [CrossRef] [PubMed] [Google Scholar]
- Duan YL, Bellis G, Li L, Li HC, Miao HS, Kou ML, Liao F, Wang Z, Gao L, Li JZ. 2019. Potential vectors of bluetongue virus in high altitude areas of Yunnan Province, China. Parasites & Vectors 12(1), 464. [CrossRef] [PubMed] [Google Scholar]
- Duan YL, Bellis G, Liu BG, Li L. 2022. Diversity and seasonal abundance of Culicoides (Diptera, Ceratopogonidae) in Shizong County, Yunnan Province, China. Parasite 29, 26. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Duan YL, Bellis G, Yang ZX, Li ZH, Liu BG, Li L 2022 DNA barcoding and phylogenetic analysis of midges belonging to Culicoides (Diptera: Ceratopogonidae) subgenus Hoffmania in Yunnan, China. Journal of Asia-Pacific Entomology 25(4), 101989. [CrossRef] [Google Scholar]
- Duan YL, Li L, Bellis G, Yang ZX, Li HC. 2021. Detection of bluetongue virus in Culicoides spp. in southern Yunnan Province, China. Parasites & Vectors 14(1), 68. [CrossRef] [PubMed] [Google Scholar]
- Duan YL, Miao HS, Liao F, Kou ML, Li ZH, Wang Z, Li HC, Li L. 2019. The serologic investigation and viral isolation of bluetongue virus in Shangri-La in Southwest China. Transboundary and Emerging Diseases 66(6), 2353–2361. [CrossRef] [PubMed] [Google Scholar]
- Duan YL, Yang ZX, Bellis G, Li L. 2021. Isolation of Tibet Orbivirus from Culicoides jacobsoni (Diptera, Ceratopogonidae) in China. Parasites & Vectors 14(1), 432. [CrossRef] [PubMed] [Google Scholar]
- Dyce AL. 1969. The recognition of nulliparous and parous Culicoides (Diptera: Ceratopogonidae) without dissection. Australian Journal of Entomology 8(1), 11–15. [CrossRef] [Google Scholar]
- Foxi C, Delrio G, Falchi G, Marche MG, Satta G, Ruiu L. 2016. Role of different Culicoides vectors (Diptera: Ceratopogonidae) in bluetongue virus transmission and overwintering in Sardinia (Italy). Parasites & Vectors 9(1), 440. [Google Scholar]
- Foxi C, Meloni G, Puggioni G, Manunta D, Rocchigiani A, Vento L, Cabras P, Satta G. 2019. Bluetongue virus detection in new Culicoides species in Sardinia, Italy. Veterinary Record 184(20), 621. [CrossRef] [PubMed] [Google Scholar]
- Fujisawa Y, Homat T, Thepparat A, Changbunjong T, Sutummaporn K, Kornmatitsuk S, Kornmatitsuk B. 2021. DNA barcode identification and molecular detection of bluetongue virus in Culicoides biting midges (Diptera: Ceratopogonidae) from western Thailand. Acta Tropica 224, 106147. [CrossRef] [PubMed] [Google Scholar]
- Hall BG. 2013. Building phylogenetic trees from molecular data with MEGA. Molecular Biology Evolution 30(5), 1229–1235. [CrossRef] [PubMed] [Google Scholar]
- He Y, Meng J, Li N, Li Z, Wang D, Kou M, Yang Z, Li Y, Zhang L, Wang J. 2024. Isolation of Epizootic Hemorrhagic Disease Virus serotype 10 from Culicoides tainanus and associated infections in livestock in Yunnan, China. Viruses 16(2), 175. [CrossRef] [PubMed] [Google Scholar]
- Hofmann M, Griot C, Chaignat V, Perler L, Thur B. 2008. Blauzungenkrankheit erreicht die Schweiz. Schweizer Archiv fur Tierheilkunde 150(2), 49–56. [CrossRef] [PubMed] [Google Scholar]
- ICTV. ICTV Master Species List 2022 MSL38.v3. Master Species Lists 2022. Available from: https://ictv.global/msl [cited 28 Feb 2024]. [Google Scholar]
- Kim HC, Bellis GA, Kim MS, Klein TA, Gopurenko D, Cai DC, Seo HJ, Cho IS, Park JY. 2015. Species diversity and seasonal distribution of Culicoides spp. (Diptera: Ceratopogonidae) in Jeju-do, Republic of Korea. Korean Journal of Parasitology 53(4), 501–506. [CrossRef] [PubMed] [Google Scholar]
- Kim MS, Kim HC, Bellis GA, Chong ST, Kim HS, Klein TA. 2021.Seasonal abundance of Culicoides at Yongsan US army garrison (USAG) and Camp Humphreys USAG, Republic of Korea, 2010–2013 and 2014–2017, Korean Journal of Parasitology 59(3), 273–280. [CrossRef] [PubMed] [Google Scholar]
- Kitaoka S, Morii T 1963. Observations on the breeding habitats of some biting midges and seasonal population dynamics in the life cycle of Culicoides arakawae in Tokyo and its vicinity. National Institute of Animal Health Quarterly 3(1), 198–208. [Google Scholar]
- Li M, Zheng Y, Zhao G, Fu S, Wang D, Wang Z, Liang G. 2014. Tibet Orbivirus, a novel Orbivirus species isolated from Anopheles maculatus mosquitoes in Tibet, China. PLoS One 9(2), e88738. [CrossRef] [PubMed] [Google Scholar]
- Li N, Meng J, He Y, Wang W, Wang J. 2023. Potential roles of Culicoides spp. (Culicoides imicola, Culicoides oxystoma) as biological vectors of bluetongue virus in Yuanyang of Yunnan, PR China. Frontiers in Cellular and Infection Microbiology 13, 1283216. [Google Scholar]
- Liu GP, Deng CY. 2000. A new Species of Culicoides and newly found male of Culicoides pelius Liu et Yu (Diptera: Ceratopogonidae) from Tibet, China. Chinese Journal of Vector Biology and Control 11(4), 245–247. [Google Scholar]
- Liu GP, Liang GD. 2021. An investigation of geographical distribution of hematophagous midges (Diptera: Ceratopogonidae) in 31 counties (banners and cities) in 8 provinces (autonomous regions) of China. Chinese Journal of Vector Biology and Control 32(4), 487–493. [Google Scholar]
- Lysyk TJ, Danyk T. 2007. Effect of temperature on life history parameters of adult Culicoides sonorensis (Diptera: Ceratopogonidae) in relation to geographic origin and vectorial capacity for bluetongue virus. Journal of Medical Entomology 44(5), 741–751. [CrossRef] [PubMed] [Google Scholar]
- Maclachlan NJ. 2011. Bluetongue: history, global epidemiology, and pathogenesis. Preventive Veterinary Medicine 102(2), 107–111. [CrossRef] [PubMed] [Google Scholar]
- Martinez-de la Puente J, Mathieu B, Carpenter S, Baldet T. 2021. Culicoides imicola (Biting Midge). Trends in Parasitology 37(5), 458–459. [CrossRef] [PubMed] [Google Scholar]
- Matsumoto Y, Yanase T, Tsuda T, Noda H. 2009. Species-specific mitochondrial gene rearrangements in biting midges and vector species identification. Medical and Veterinary Entomology 23(1), 47–55. [CrossRef] [PubMed] [Google Scholar]
- Meiswinkel R, Gomulski LM, Delecolle JC, Goffredo M, Gasperi G. 2004. The taxonomy of Culicoides vector complexes – unfinished business, Veterinaria Italiana 40(3), 151–159. [PubMed] [Google Scholar]
- Mellor PS, Boorman J, Baylis M. 2000. Culicoides biting midges: their role as arbovirus vectors. Annual Review of Entomology 45(1), 307–340. [CrossRef] [PubMed] [Google Scholar]
- Mendez-Lopez MR, Attoui H, Florin D, Calisher CH, Florian-Carrillo JC, Montero S. 2015. Association of vectors and environmental conditions during the emergence of Peruvian horse sickness orbivirus and Yunnan orbivirus in northern Peru. Journal of Vector Ecology 40(2), 355–363. [CrossRef] [PubMed] [Google Scholar]
- Napp S, Gubbins S, Calistri P, Allepuz A, Alba A, Garcia-Bocanegra I, Giovannini A, Casal J. 2011. Quantitative assessment of the probability of bluetongue virus overwintering by horizontal transmission: application to Germany. Veterinary Research 42(1), 4. [CrossRef] [PubMed] [Google Scholar]
- OIE. Epizootic haemorrhagic disease (infection with epizootic hemorrhagic disease virus). OIE Terrestrial Manual 2021. Available from: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.07_EHD.pdf [cited 2021 30 Nov 2021]. [Google Scholar]
- Omori T, Inaba Y, Morimoto T, Tanaka Y, Ishitani R. 1969. Ibaraki virus, an agent of epizootic disease of cattle resembling bluetongue. I. Epidemiologic, clinical and pathologic observations and experimental transmission to calves. Japanese Journal of Microbiology 13(2), 139–157. [CrossRef] [PubMed] [Google Scholar]
- Osborne CJ, Mayo CE, Mullens BA, McDermott EG, Gerry AC, Reisen WK, MacLachlan NJ. 2015. Lack of evidence for laboratory and natural vertical transmission of Bluetongue Virus in Culicoides sonorensis (Diptera: Ceratopogonidae). Journal of Medical Entomology 52(2), 274–277. [CrossRef] [PubMed] [Google Scholar]
- Peng HH. 2020. Identification of new mosquito-borne viruses and their molecular characteristics and serological survey for the virus infections in Yunnan Province. Hefei: Anhui Medical University. [Google Scholar]
- Peng HH, Wang YY, Dai K, Zhao QM, Zuo SQ, Tong YG, Zhang JS. 2020. Molecular characterization of two variant totiviruses isolated from Culex tritaeniorhynchus in China. Acta Parasitologica et Medica Entomologica Sinica 27(1), 20–25. [Google Scholar]
- Purse BV, Carpenter S, Venter GJ, Bellis G, Mullens BA. 2015. Bionomics of temperate and tropical Culicoides midges: knowledge gaps and consequences for transmission of Culicoides-borne viruses. Annual Review of Entomology 60, 373–392. [CrossRef] [PubMed] [Google Scholar]
- Purse BV, Mellor PS, Rogers DJ, Samuel AR, Mertens PP, Baylis M. 2005. Climate change and the recent emergence of bluetongue in Europe. Nature Review Microbiology 3(2), 171–181. [CrossRef] [PubMed] [Google Scholar]
- Roy P. 2007. Orbiviruses, in: Fields virology, Knipe DM, Howley PM, Editors. Wolters Kluwer/Lippincott Williams & Wilkins Health: Philadelphia, PA. p. 2541–2568. [Google Scholar]
- Saminathan M, Singh KP, Khorajiya JH, Dinesh M, Vineetha S, Maity M, Rahman AF, Misri J, Malik YS, Gupta VK, Singh RK, Dhama K. 2020. An updated review on bluetongue virus: epidemiology, pathobiology, and advances in diagnosis and control with special reference to India. Veterinary Quarterly 40(1), 258–321. [CrossRef] [PubMed] [Google Scholar]
- Standfast HA, Dyce AL, Muller MJ. 1985. Vectors of bluetongue virus in Australia, in: Bluetongue and related orbiviruses, Barber TL, Jochim MM, Editors. Liss: New York, p. 177–186. [Google Scholar]
- Steyn J, Venter GJ, Labuschagne K, Majatladi D, Boikanyo SN, Lourens C, Ebersohn K, Venter EH. 2016. Possible over-wintering of bluetongue virus in Culicoides populations in the Onderstepoort area, Gauteng, South Africa. Journal of the South African Veterinary Association 87(1), e1–e5. [CrossRef] [PubMed] [Google Scholar]
- Takamatsu H, Mellor PS, Mertens PPC, Kirkham PA, Burroughs JN, Parkhouse RME. 2003. A possible overwintering mechanism for bluetongue virus in the absence of the insect vector. Journal of General Virology 84(Pt 1), 227–235. [CrossRef] [PubMed] [Google Scholar]
- Van der Saag M, Nicholas A, Ward M, Kirkland P. 2015. Evaluation of in vitro methods for assessment of infection of Australian Culicoides spp. with bluetongue viruses. Veterinaria Italiana 51(4), 335–343. [PubMed] [Google Scholar]
- Van Dijk AA, Huismans H. 1982. The effect of temperature on the in vitro transcriptase reaction of bluetongue virus, epizootic haemorrhagic disease virus and African horsesickness virus. Onderstepoort Journal of Veterinary Research 49(4), 227–232. [Google Scholar]
- Veronesi E, Antony F, Gubbins S, Golding N, Blackwell A, Mertens PP, Brownlie J, Darpel KE, Mellor PS, Carpenter S. 2013. Measurement of the infection and dissemination of bluetongue virus in Culicoides biting midges using a semi-quantitative RT-PCR assay and isolation of infectious virus. PLoS One 8(8), e70800. [CrossRef] [PubMed] [Google Scholar]
- Viadanna PHO, Rodrigues TCS, Subramaniam K, Campos Krauer JM, Lednicky JA, Loeb JC, Wisely SM, Waltzek TB. 2021. Genome sequence of a Yunnan Orbivirus isolated from a dead Florida White-Tailed Deer (Odocoileus virginianus). Microbiology Resource Announcements 10(18), e00168-21. [CrossRef] [PubMed] [Google Scholar]
- Wang J, Li H, He Y, Zhou Y, Xin A, Liao D, Meng J. 2017. Isolation of Tibet Orbivirus from Culicoides and associated infections in livestock in Yunnan, China. Virology Journal 14(1), 105. [CrossRef] [PubMed] [Google Scholar]
- White DM, Wilson WC, Blair CD, Beaty BJ. 2005. Studies on overwintering of bluetongue viruses in insects. Journal of General Virology 86(Pt 2), 453–462. [CrossRef] [PubMed] [Google Scholar]
- Wickner RB. 2007. Viruses and prions of yeasts, fungi, and unicellular organisms, in: Fields virology, Knipe DM, Howley PM, Editors. Wolters Kluwer/Lippincott Williams & Wilkins Health: Philadelphia, PA, p. 738–769. [Google Scholar]
- Wirth WW, Hubert AA. 1989. The Culicoides of Southeast Asia (Diptera: Ceratopogonidae). Memoirs of the American Entomological Institute 44(1), 1–509. [Google Scholar]
- Yanase T, Murota K, Hayama Y. 2020. Endemic and emerging arboviruses in domestic ruminants in East Asia. Frontiers in Veterinary Science 7, 168. [CrossRef] [PubMed] [Google Scholar]
- Yang H, Xiao L, Wang J, Meng J, Lv M, Liao D, Song J, Gao L, Xiong H, He Y, Niu B, Chuang X, Li H. 2017. Phylogenetic characterization genome segment 2 of Bluetongue virus strains belonging to serotypes 5, 7 and 24 isolated for the first time in China during 2012 to 2014, Transboundary and Emerging Diseases 64(4), 1317–1321. [CrossRef] [PubMed] [Google Scholar]
- Yang X, Zhang Y, Ge X, Yuan J, Shi Z. 2012. A novel totivirus-like virus isolated from bat guano. Archives of Virology 157(6), 1093–1099. [CrossRef] [PubMed] [Google Scholar]
- Yang ZX, Zhu JB, Li ZH, Li ZR, He YW, Xie JR, Li HC, Liao DF, Yang H. 2019. Establishment and application of a duplex real-time RT-PCR assay for simultaneous detection of bluetongue virus and epidemic haemorrhagic disease virus. Chinese Veterinary Science 49(9), 1104–1111. [Google Scholar]
- Yu YX, Liu JH, Liu GP, Liu ZJ, Yan G, Hao BS, Zhao TS, Yu YX, . 2005. Ceratopogonidae: Culicoides, in: Ceratopogonidae of China, Yu YX, Editor. Military Medical Science Press: Beijing, p. 816–1323. [Google Scholar]
- Zhang NZ, Zhang KL, Li ZH, Chan-yu NC, Hu YL, Li G, Zhao K, Zou FZ, Xu WZ, Li SX, Li CD, Zhang YS, Xu YS, Liu SH, Zhou XC, Dou WX, Bao CH, Zhao XQ. 1989. A report of the investigation and research of Bluetongue on sheep. Yunnan Journal of Animal Science and Veterinary Medicine 18(4), 3–13. [Google Scholar]
Cite this article as: Duan Y-L, Li Z-H, Bellis GA, Li L, Liu B-G,Wang J-P, Liu J-M, Liao D-F & Zhu J-B. 2024. Culicoides and midgeassociated arboviruses on cattle farms in Yunnan Province, China. Parasite 31, 72. https://doi.org/10.1051/parasite/2024072.
All Tables
Diversity and abundance of Culicoides collected over a 12-month period from 2 farms in Lufeng County, Yunnan Province, China.
Proportion of male specimens in UV trap collections of Culicoides collected on 2 farms over 12 months in Lufeng County, Yunnan Province, China.
Viruses isolated from pools of Culicoides and cattle blood collected from 2 farms in Lufeng County, Yunnan Province, China.
RT-qPCR results from pools of parous + gravid Culicoides collected in Lufeng County, Yunnan Province, China.
All Figures
|
Figure 1 Wings of represented female specimens. The wings of represented Culicoides species collected from Lufeng County in this study, scale bar = 0.5 mm. |
| In the text | |
|
Figure 2 Abundance of Culicoides species on two farms in Lufeng Province, Yunnan, China. Each cell represents a collection and the color reflects the Culicoides abundance in each collection. |
| In the text | |
|
Figure 3 Seasonal abundance of Culicoides and weather on 2 farms in Lufeng County, Yunnan, China. (A) The relative proportion of female midges belonging to the major species present in collections from Mar 2022 to Mar 2023. (B) Total midge abundance (log10 (n + 1)) on Farm A and Farm B, and the average abundance at the two farms from Mar 2022 to Mar 2023. (C) Daily maximum and minimum temperature (TEM) and rain score between March 1, 2022 and March 31, 2023 in Lufeng County. |
| In the text | |
|
Figure 4 Proportions of blood fed and nulliparous females. The average percentages (mean ± SD) of blood feeding (A) and nulliparous female (B) Culicoides collected on 2 farms in Lufeng county, Yunnan, China between April and December 2022. |
| In the text | |
|
Figure 5 Phylogenetic analysis for YUOV and YSToV, and related viruses. Phylogenetic trees of T2 genes of YUOV, MPOV, and BTV (A), and RdRP genes of YSToV and close totivirus (B) were constructed by NJ algorithm. The sequences of our isolates (contig-214, contig-S5) were marked by circles. Bootstrap values less than 70% were omitted. |
| In the text | |
|
Figure S1: Locality map of Yunnan and flow chart of experiment. (A) Sketch map of Yunnan Province, China. Major rivers and mountains are shown roughly by green and grey lines, respectively. (B) Collecting samples on Farm A. (C) Preliminary RT-qPCR test for Culicoides. (D) Further RT-qPCR test for positive samples. |
| In the text | |
|
Figure S2: Identification of viral isolates by RT-PCR. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.